

Evolution of deep gray matter volume across the human lifespan
- PMID: 28548250
- PMCID: PMC6867004
- DOI: 10.1002/hbm.23604
Evolution of deep gray matter volume across the human lifespan
Abstract
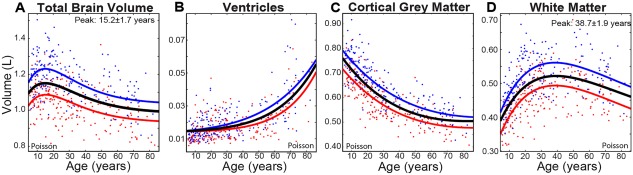
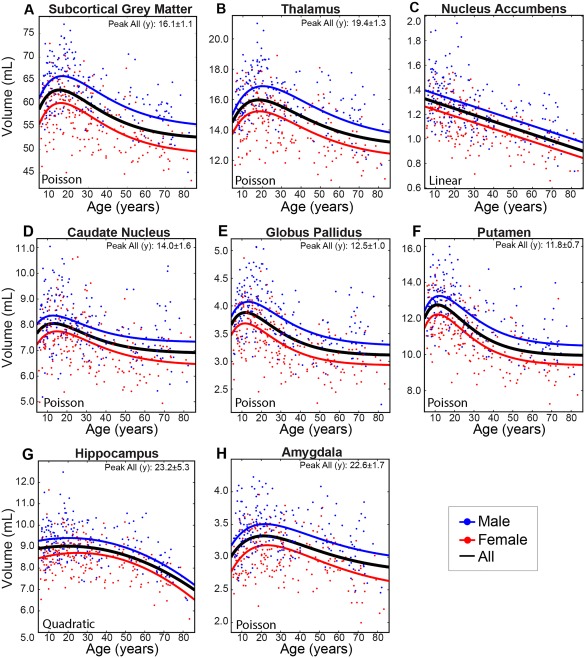
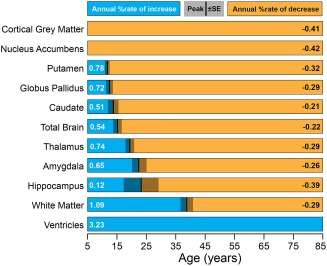
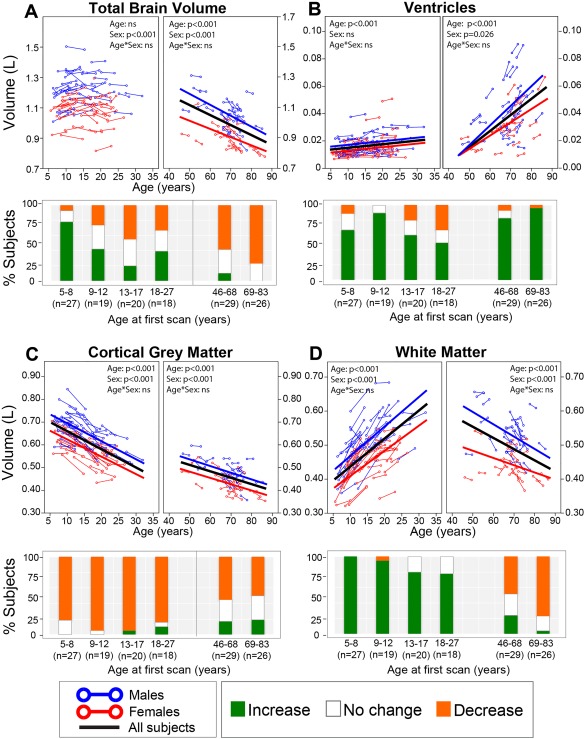
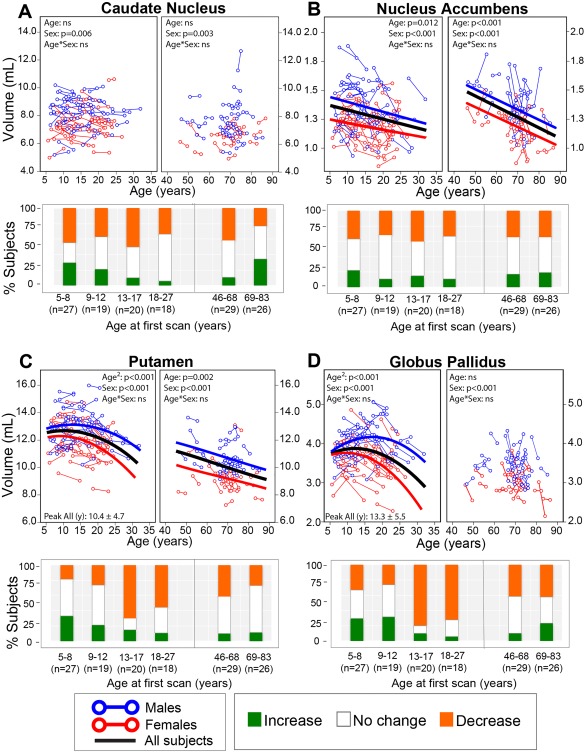
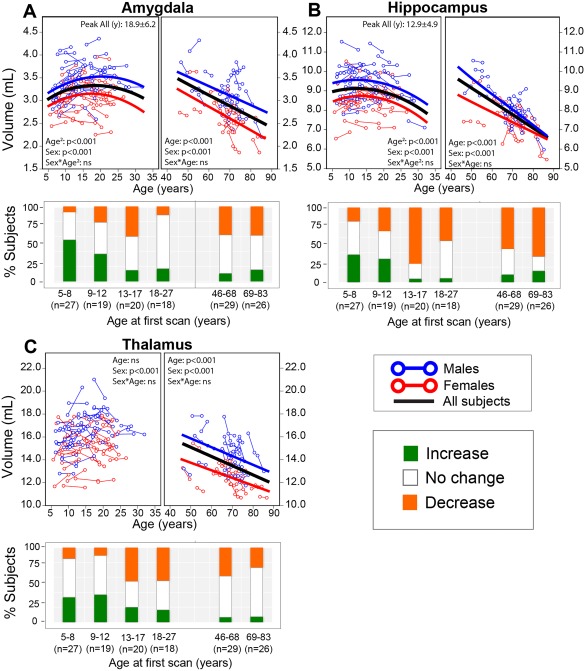
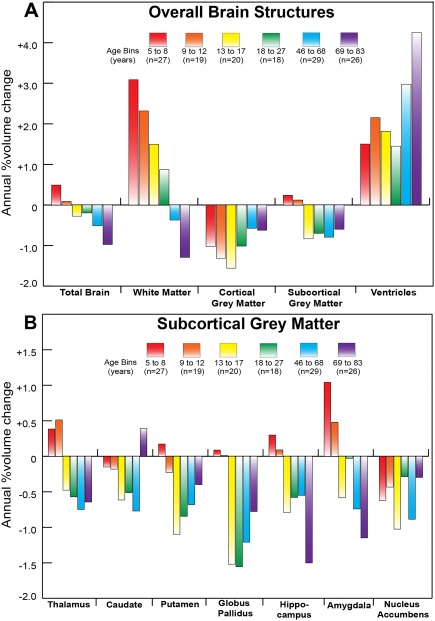
Magnetic resonance imaging of subcortical gray matter structures, which mediate behavior, cognition and the pathophysiology of several diseases, is crucial for establishing typical maturation patterns across the human lifespan. This single site study examines T1-weighted MPRAGE images of 3 healthy cohorts: (i) a cross-sectional cohort of 406 subjects aged 5-83 years; (ii) a longitudinal neurodevelopment cohort of 84 subjects scanned twice approximately 4 years apart, aged 5-27 years at first scan; and (iii) a longitudinal aging cohort of 55 subjects scanned twice approximately 3 years apart, aged 46-83 years at first scan. First scans from longitudinal subjects were included in the cross-sectional analysis. Age-dependent changes in thalamus, caudate, putamen, globus pallidus, nucleus accumbens, hippocampus, and amygdala volumes were tested with Poisson, quadratic, and linear models in the cross-sectional cohort, and quadratic and linear models in the longitudinal cohorts. Most deep gray matter structures best fit to Poisson regressions in the cross-sectional cohort and quadratic curves in the young longitudinal cohort, whereas the volume of all structures except the caudate and globus pallidus decreased linearly in the longitudinal aging cohort. Males had larger volumes than females for all subcortical structures, but sex differences in trajectories of change with age were not significant. Within subject analysis showed that 65%-80% of 13-17 year olds underwent a longitudinal decrease in volume between scans (∼4 years apart) for the putamen, globus pallidus, and hippocampus, suggesting unique developmental processes during adolescence. This lifespan study of healthy participants will form a basis for comparison to neurological and psychiatric disorders. Hum Brain Mapp 38:3771-3790, 2017. © 2017 Wiley Periodicals, Inc.
Keywords: FreeSurfer; Poisson curve; aging; basal ganglia; limbic structures; neurodevelopment; subcortical gray matter; thalamus.
© 2017 Wiley Periodicals, Inc.
Figures
References
-
- Bouchard TP, Malykhin N, Martin WR, Hanstock CC, Emery DJ, Fisher NJ, Camicioli RM (2008): Age and dementia‐associated atrophy predominates in the hippocampal head and amygdala in Parkinson's disease. Neurobiol Aging 29:1027–1039. - PubMed
-
- Brown TT, Kuperman JM, Chung YH, Erhart M, McCabe C, Hagler DJ, Venkatraman VK, Akshoomoff N, Amaral DG, Bloss CS, Casey BJ, Chang L, Ernst TM, Frazier JA, Gruen JR, Kaufmann WE, Kenet T, Kennedy DN, Murray SS, Sowell ER, Jernigan TL, Dale AM (2012): Neuroanatomical assessment of biological maturity. Curr Biol 22:1693–1698. - PMC - PubMed
-
- Brown CJ, Miller SP, Booth BG, Andrews S, Chau V, Poskitt KJ, Hamarneh G (2014): Structural network analysis of brain development in young preterm neonates. Neuroimage 101:667–680. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
