

Nicotinamide Adenine Dinucleotide Metabolism and Neurodegeneration
- PMID: 28548540
- PMCID: PMC5962335
- DOI: 10.1089/ars.2017.7145
Nicotinamide Adenine Dinucleotide Metabolism and Neurodegeneration
Abstract
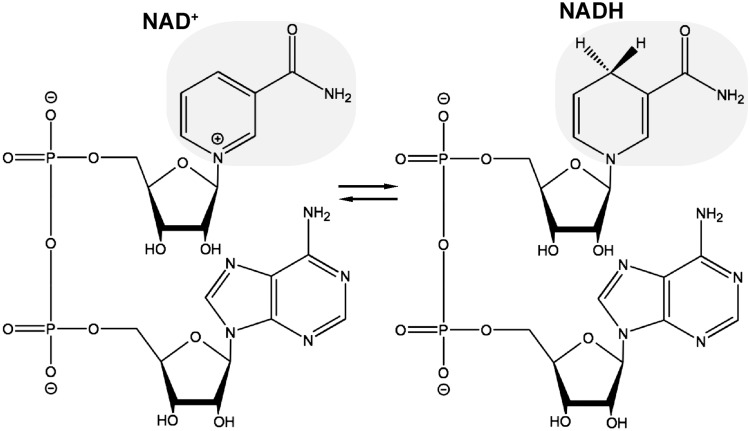
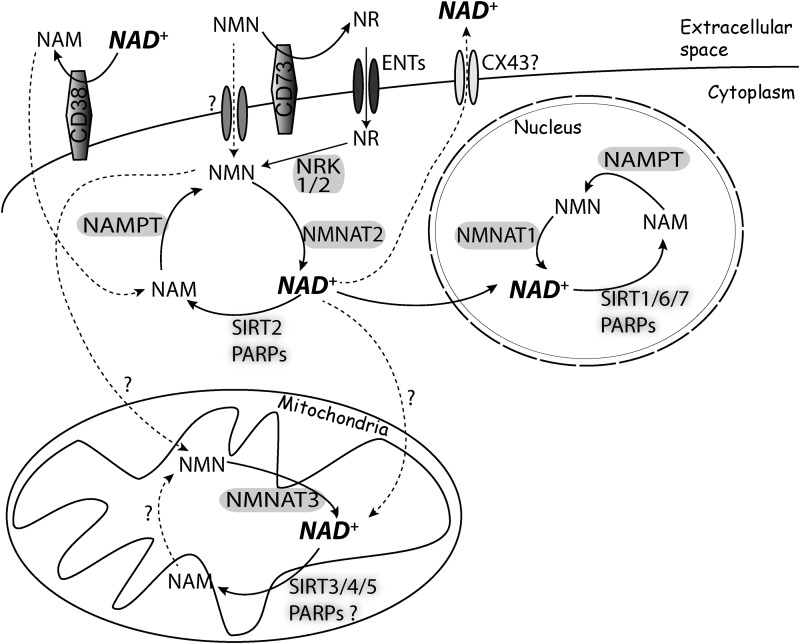
Significance: Nicotinamide adenine dinucleotide (NAD+) participates in redox reactions and NAD+-dependent signaling processes, which involve the cleavage of NAD+ coupled to posttranslational modifications of proteins or the production of second messengers. Either as a primary cause or as a secondary component of the pathogenic process, mitochondrial dysfunction and oxidative stress are prominent features of several neurodegenerative diseases. Activation of NAD+-dependent signaling pathways has a major effect in the capacity of the cell to modulate mitochondrial function and counteract the deleterious effects of increased oxidative stress. Recent Advances: Progress in the understanding of the biological functions and compartmentalization of NAD+-synthesizing and NAD+-consuming enzymes have led to the emergence of NAD+ metabolism as a major therapeutic target for age-related diseases.
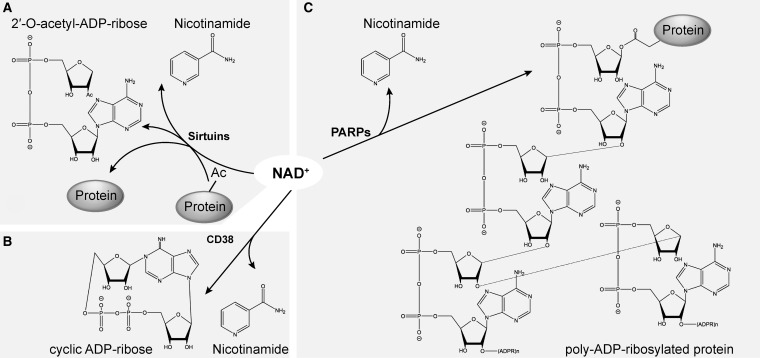
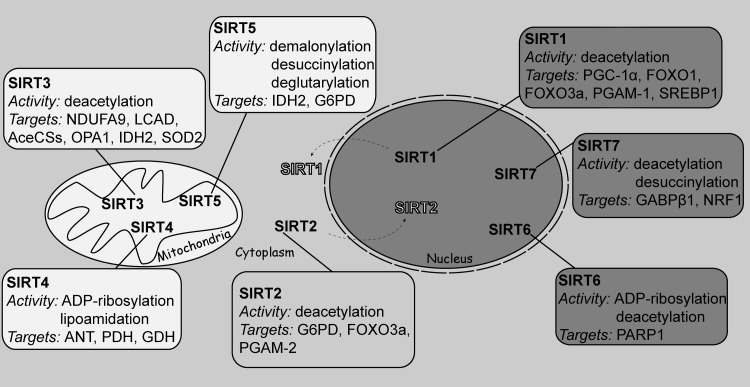
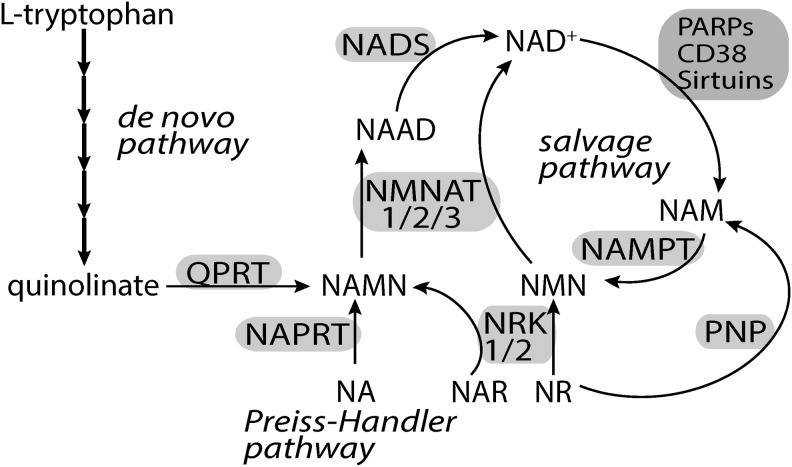
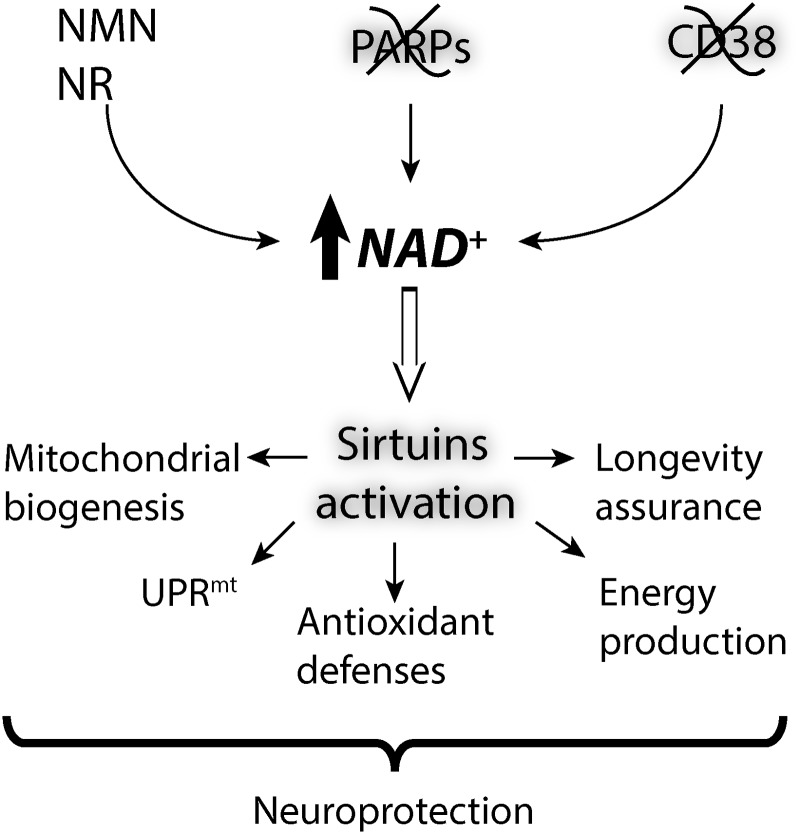
Critical issues: Three distinct families of enzymes consume NAD+ as substrate: poly(ADP-ribose) polymerases (PARPs), ADP-ribosyl cyclases (CD38/CD157) and sirtuins. Two main strategies to increase NAD+ availability have arisen. These strategies are based on the utilization of NAD+ intermediates/precursors or the inhibition of the NAD+-consuming enzymes, PARPs and CD38. An increase in endogenous sirtuin activity seems to mediate the protective effect that enhancing NAD+ availability confers in several models of neurodegeneration and age-related diseases.
Future directions: A growing body of evidence suggests the beneficial role of enhancing NAD+ availability in models of neurodegeneration. The challenge ahead is to establish the value and safety of the long-term use of these strategies for the treatment of neurodegenerative diseases. Antioxid. Redox Signal. 28, 1652-1668.
Keywords: CD38; NAD; PARP; mitochondria; neurodegeneration; oxidative stress.
Figures
References
-
- Abeti R, Abramov AY, and Duchen MR. Beta-amyloid activates PARP causing astrocytic metabolic failure and neuronal death. Brain 134: 1658–1672, 2011 - PubMed
-
- Ahuja N, Schwer B, Carobbio S, Waltregny D, North BJ, Castronovo V, Maechler P, and Verdin E. Regulation of insulin secretion by SIRT4, a mitochondrial ADP-ribosyltransferase. J Biol Chem 282: 33583–33592, 2007 - PubMed
-
- Aksoy P, Escande C, White TA, Thompson M, Soares S, Benech JC, and Chini EN. Regulation of SIRT 1 mediated NAD dependent deacetylation: a novel role for the multifunctional enzyme CD38. Biochem Biophys Res Commun 349: 353–359, 2006 - PubMed
-
- Aksoy P, White TA, Thompson M, and Chini EN. Regulation of intracellular levels of NAD: a novel role for CD38. Biochem Biophys Res Commun 345: 1386–1392, 2006 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
