

The constitutive lipid droplet protein PLIN2 regulates autophagy in liver
- PMID: 28548876
- PMCID: PMC5529083
- DOI: 10.1080/15548627.2017.1319544
The constitutive lipid droplet protein PLIN2 regulates autophagy in liver
Abstract
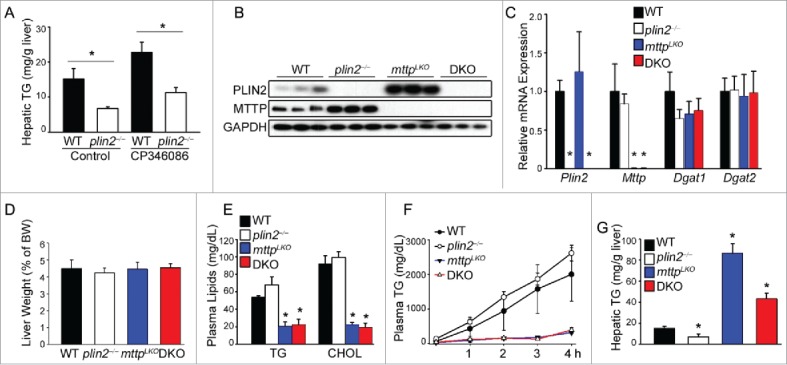
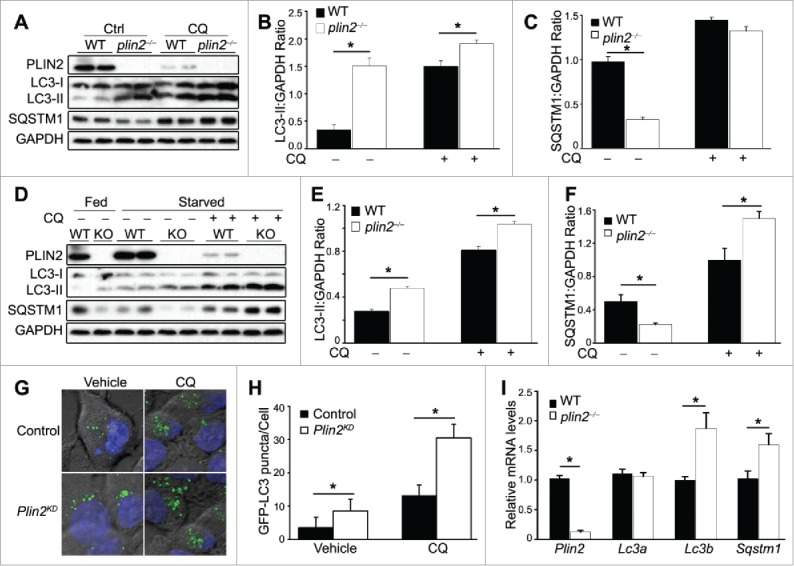
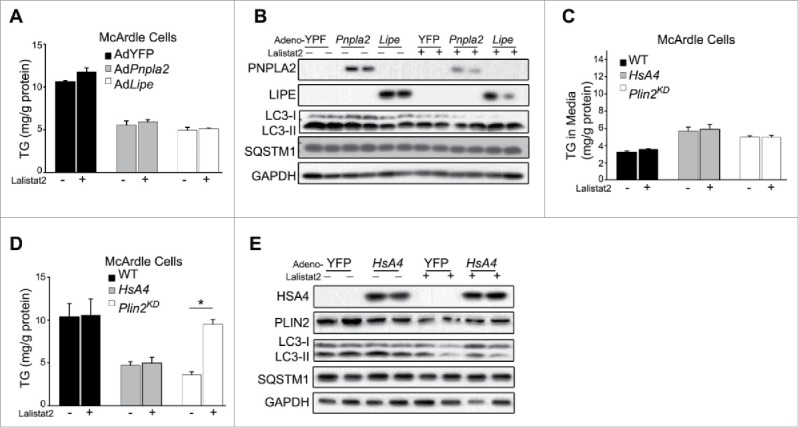
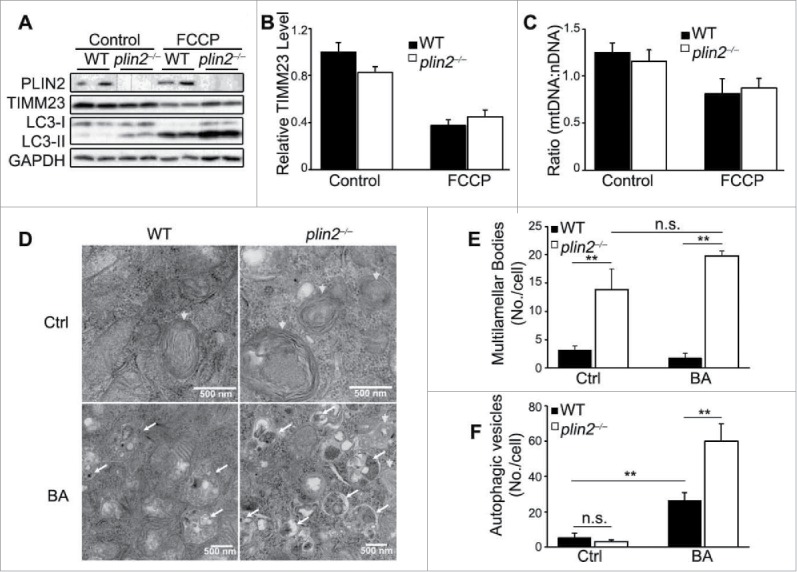
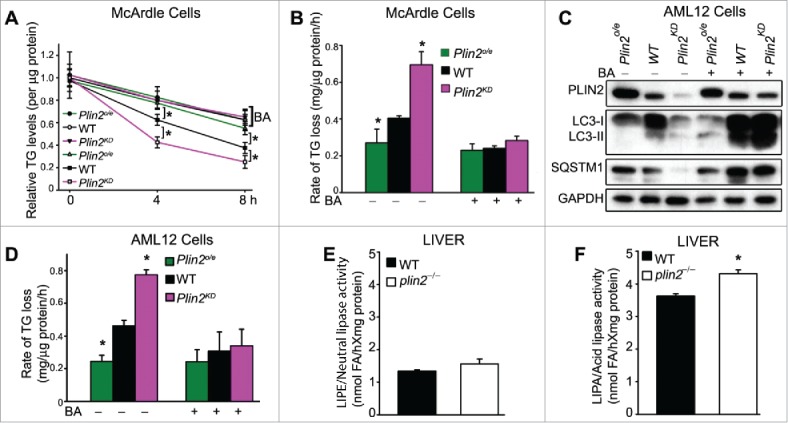
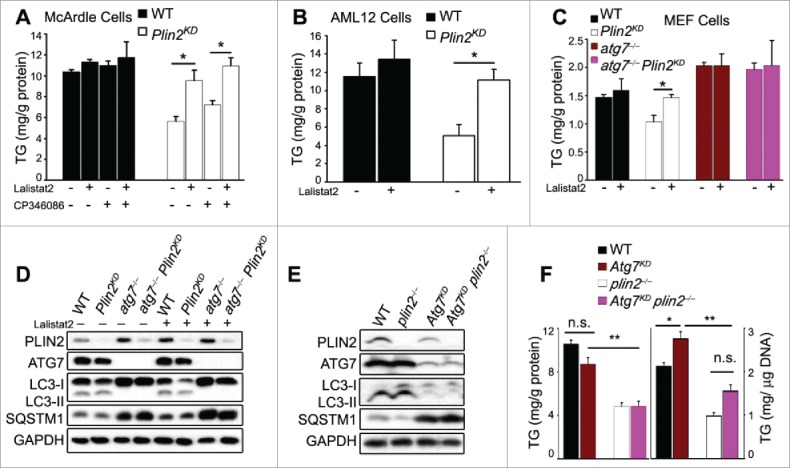
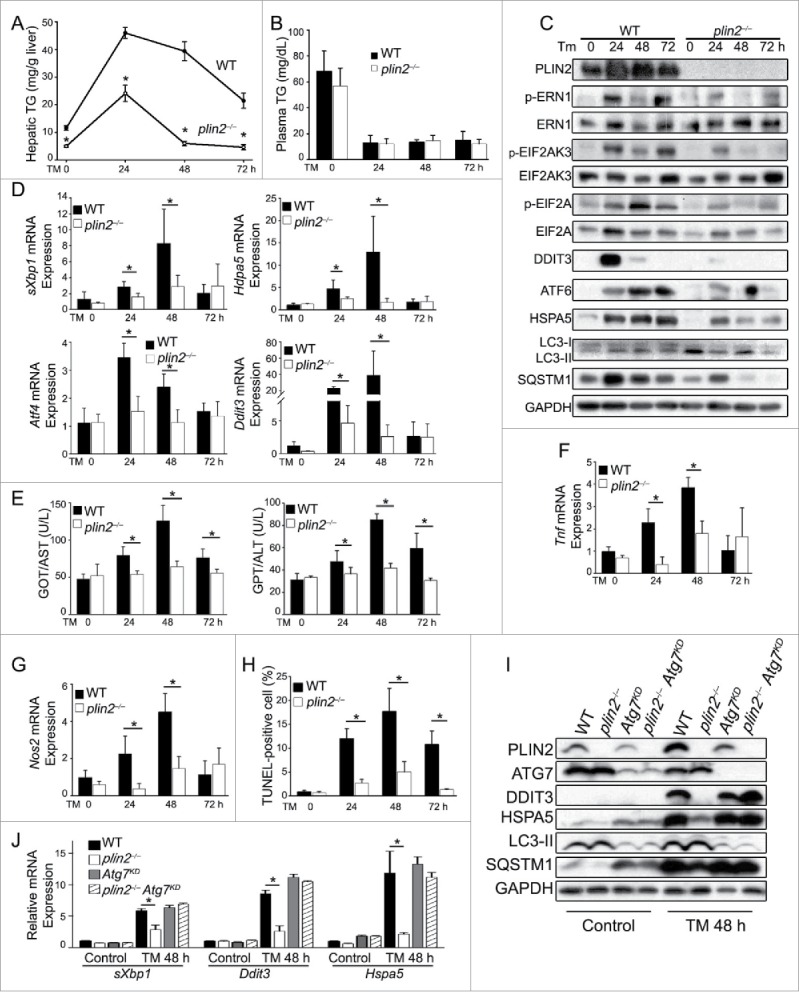
Excess triglyceride (TG) accumulation in the liver underlies fatty liver disease, a highly prevalent ailment. TG occurs in the liver sequestered in lipid droplets, the major lipid storage organelle. Lipid droplets are home to the lipid droplet proteins, the most abundant of which are the perilipins (PLINs), encoded by 5 different genes, Plin1 to Plin5. Of the corresponding gene products, PLIN2 is the only constitutive and ubiquitously expressed lipid droplet protein that has been used as a protein marker for lipid droplets. We and others reported that plin2-/- mice have an ∼60% reduction in TG content, and are protected against fatty liver disease. Here we show that PLIN2 overexpression protects lipid droplets against macroautophagy/autophagy, whereas PLIN2 deficiency enhances autophagy and depletes hepatic TG. The enhanced autophagy in plin2-/- mice protects against severe ER stress-induced hepatosteatosis and hepatocyte apoptosis. In contrast, hepatic TG depletion resulting from other genetic and pharmacological manipulations has no effect on autophagy. Importantly, PLIN2 deficiency lowers cellular TG content in wild-type mouse embryonic fibroblasts (MEFs) via enhanced autophagy, but does not affect cellular TG content in atg7-/- MEFs that are devoid of autophagic function. Conversely, adenovirus-shAtg7-mediated hepatic Atg7 knockdown per se does not alter the hepatic TG level, suggesting a more complex regulation in vivo. In sum, PLIN2 guards its own house, the lipid droplet. PLIN2 overexpression protects against autophagy, and its downregulation stimulates TG catabolism via autophagy.
Keywords: ADRP; PLIN2; autophagy; hepatic TG; lipophagy; neutral lipase.
Figures
References
-
- Kawano Y, Cohen DE. Mechanisms of hepatic triglyceride accumulation in non-alcoholic fatty liver disease. J Gastroenterol 2013; 48(4):434-41; https://doi.org/ 10.1007/s00535-013-0758-5 - DOI - PMC - PubMed
-
- Quiroga AD, Lehner R. Liver triacylglycerol lipases. Biochim Biophys Acta 2012; 1821(5):762-9; https://doi.org/ 10.1016/j.bbalip.2011.09.007 - DOI - PubMed
-
- Sekiya M, Osuga J, Yahagi N, Okazaki H, Tamura Y, Igarashi M, Takase S, Harada K, Okazaki S, Iizuka Y, et al.. Hormone-sensitive lipase is involved in hepatic cholesteryl ester hydrolysis. J Lipid Res 2008; 49(8):1829-38; https://doi.org/ 10.1194/jlr.M800198-JLR200 - DOI - PubMed
-
- Wu JW, Wang SP, Alvarez F, Casavant S, Gauthier N, Abed L, Soni KG, Yang G, Mitchell GA. Deficiency of liver adipose triglyceride lipase in mice causes progressive hepatic steatosis. Hepatology 2011; 54(1):122-32; https://doi.org/ 10.1002/hep.24338 - DOI - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous