

Regulatory T Cells in Skin Facilitate Epithelial Stem Cell Differentiation
- PMID: 28552347
- PMCID: PMC5504703
- DOI: 10.1016/j.cell.2017.05.002
Regulatory T Cells in Skin Facilitate Epithelial Stem Cell Differentiation
Abstract
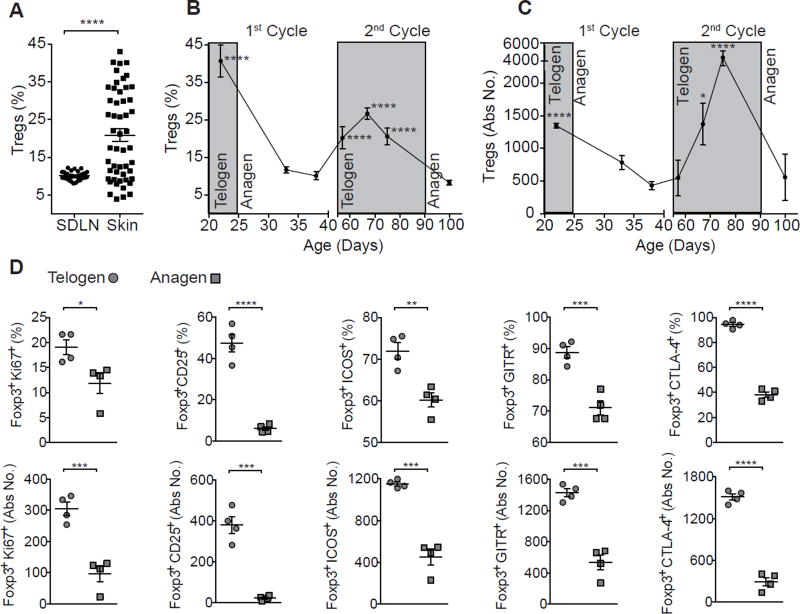
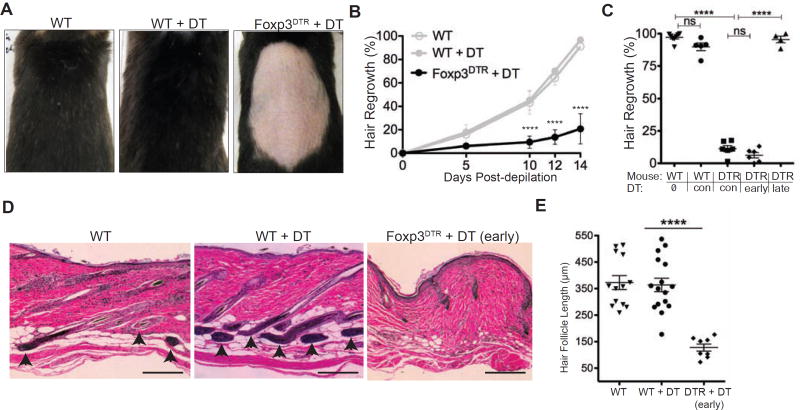
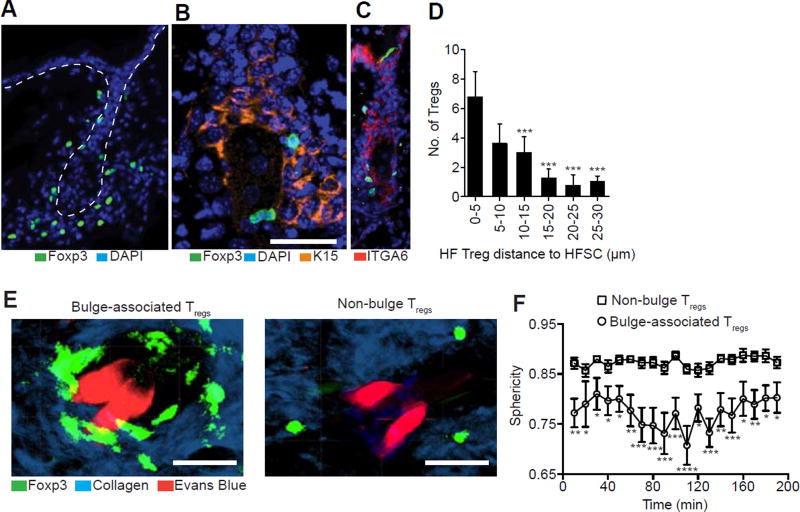
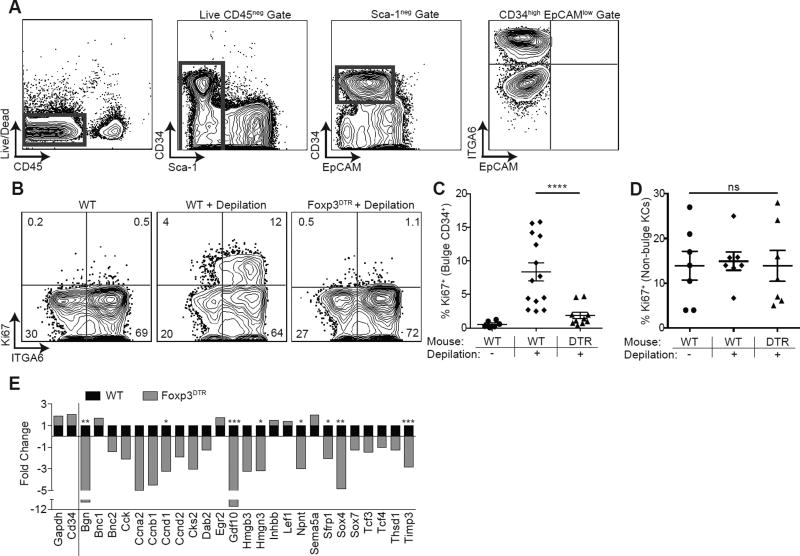
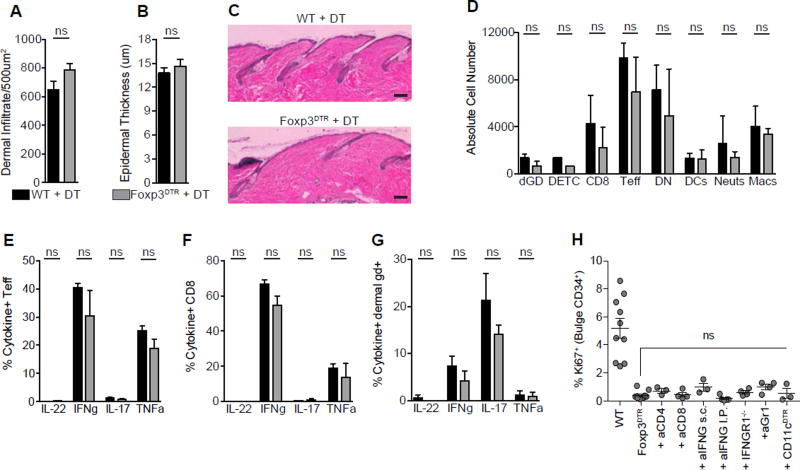
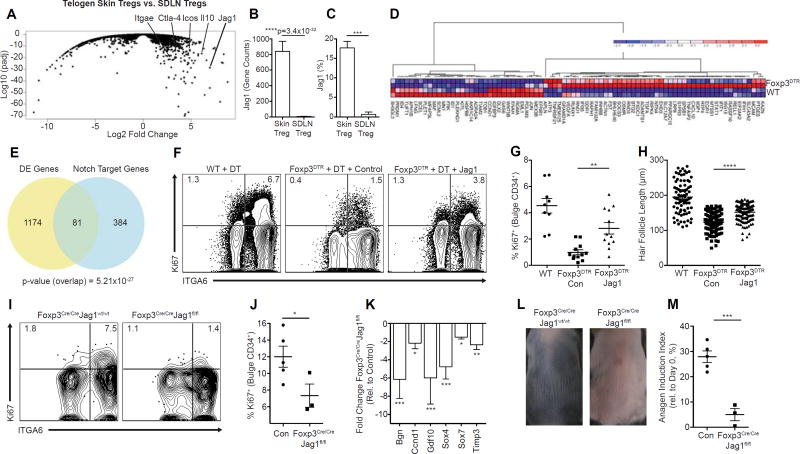
The maintenance of tissue homeostasis is critically dependent on the function of tissue-resident immune cells and the differentiation capacity of tissue-resident stem cells (SCs). How immune cells influence the function of SCs is largely unknown. Regulatory T cells (Tregs) in skin preferentially localize to hair follicles (HFs), which house a major subset of skin SCs (HFSCs). Here, we mechanistically dissect the role of Tregs in HF and HFSC biology. Lineage-specific cell depletion revealed that Tregs promote HF regeneration by augmenting HFSC proliferation and differentiation. Transcriptional and phenotypic profiling of Tregs and HFSCs revealed that skin-resident Tregs preferentially express high levels of the Notch ligand family member, Jagged 1 (Jag1). Expression of Jag1 on Tregs facilitated HFSC function and efficient HF regeneration. Taken together, our work demonstrates that Tregs in skin play a major role in HF biology by promoting the function of HFSCs.
Keywords: Jagged 1; Notch; alopecia areata; hair; hair follicle stem cell; hair regeneration; regulatory T cell; skin.
Copyright © 2017 Elsevier Inc. All rights reserved.
Figures
Comment in
-
Tregs Expand the Skin Stem Cell Niche.Dev Cell. 2017 Jun 5;41(5):455-456. doi: 10.1016/j.devcel.2017.05.020. Dev Cell. 2017. PMID: 28586641
-
T-Regulating Hair Follicle Stem Cells.Immunity. 2017 Jun 20;46(6):979-981. doi: 10.1016/j.immuni.2017.06.011. Immunity. 2017. PMID: 28636968
-
Regulatory T cells: Keep your hair on.Nat Rev Immunol. 2017 Jun 27;17(7):402-403. doi: 10.1038/nri.2017.72. Nat Rev Immunol. 2017. PMID: 28652599 No abstract available.
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
