

A Model of Fast Hebbian Spike Latency Normalization
- PMID: 28555102
- PMCID: PMC5430963
- DOI: 10.3389/fncom.2017.00033
A Model of Fast Hebbian Spike Latency Normalization
Abstract
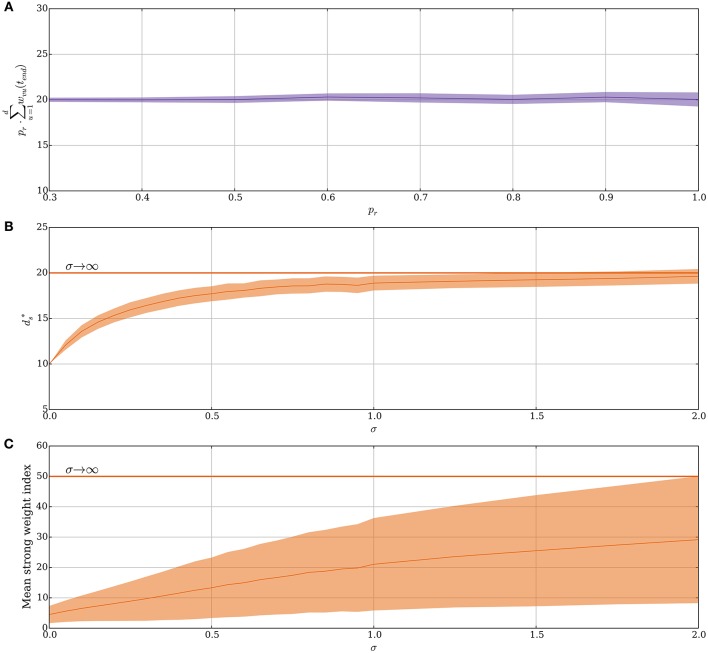
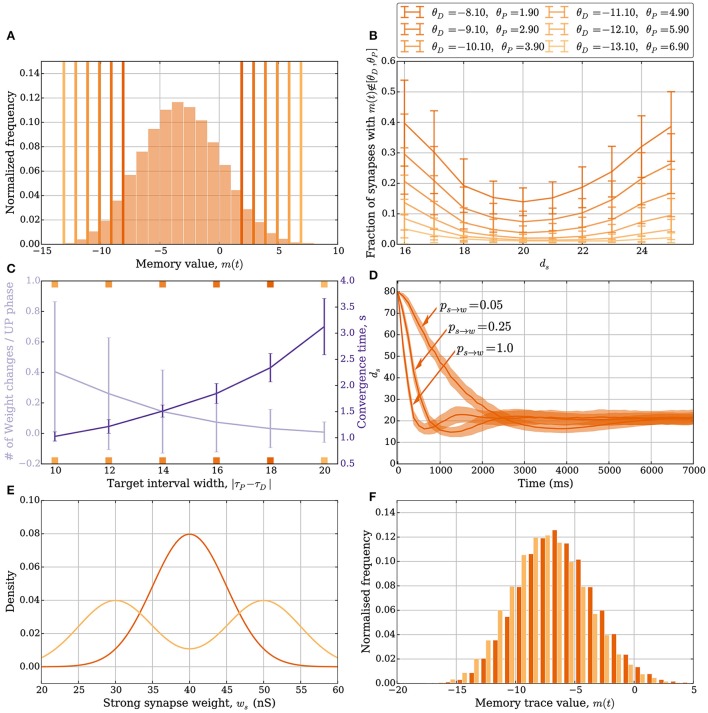
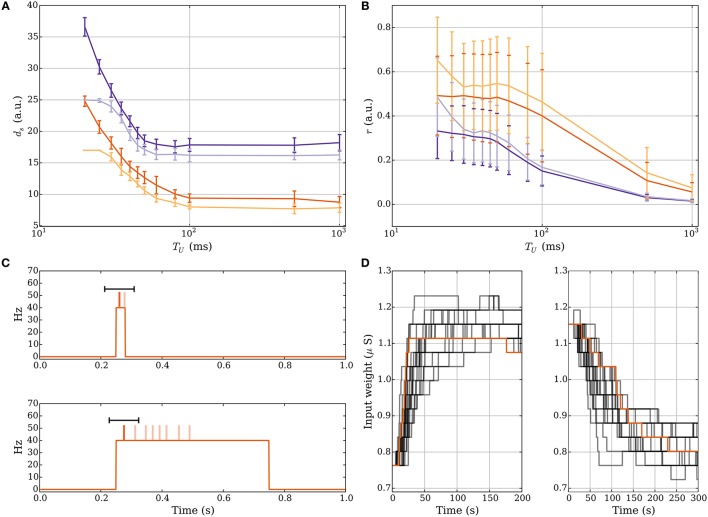
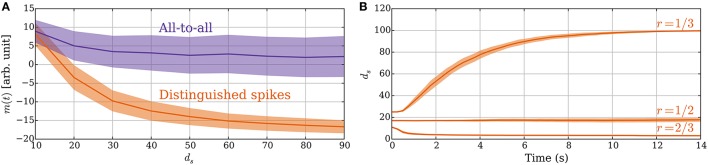
Hebbian changes of excitatory synapses are driven by and enhance correlations between pre- and postsynaptic neuronal activations, forming a positive feedback loop that can lead to instability in simulated neural networks. Because Hebbian learning may occur on time scales of seconds to minutes, it is conjectured that some form of fast stabilization of neural firing is necessary to avoid runaway of excitation, but both the theoretical underpinning and the biological implementation for such homeostatic mechanism are to be fully investigated. Supported by analytical and computational arguments, we show that a Hebbian spike-timing-dependent metaplasticity rule, accounts for inherently-stable, quick tuning of the total input weight of a single neuron in the general scenario of asynchronous neural firing characterized by UP and DOWN states of activity.
Keywords: STDP; homeostasis; metaplasticity; oscillations; synapse memory; synchrony.
Figures




Similar articles
-
Synaptic plasticity in neural networks needs homeostasis with a fast rate detector.PLoS Comput Biol. 2013;9(11):e1003330. doi: 10.1371/journal.pcbi.1003330. Epub 2013 Nov 14. PLoS Comput Biol. 2013. PMID: 24244138 Free PMC article.
-
Partial Breakdown of Input Specificity of STDP at Individual Synapses Promotes New Learning.J Neurosci. 2016 Aug 24;36(34):8842-55. doi: 10.1523/JNEUROSCI.0552-16.2016. J Neurosci. 2016. PMID: 27559167 Free PMC article.
-
The effect of STDP temporal kernel structure on the learning dynamics of single excitatory and inhibitory synapses.PLoS One. 2014 Jul 7;9(7):e101109. doi: 10.1371/journal.pone.0101109. eCollection 2014. PLoS One. 2014. PMID: 24999634 Free PMC article.
-
Models of Metaplasticity: A Review of Concepts.Front Comput Neurosci. 2015 Nov 10;9:138. doi: 10.3389/fncom.2015.00138. eCollection 2015. Front Comput Neurosci. 2015. PMID: 26617512 Free PMC article. Review.
-
Homeostatic role of heterosynaptic plasticity: models and experiments.Front Comput Neurosci. 2015 Jul 13;9:89. doi: 10.3389/fncom.2015.00089. eCollection 2015. Front Comput Neurosci. 2015. PMID: 26217218 Free PMC article. Review.
Cited by
-
Biological data questions the support of the self inhibition required for pattern generation in the half center model.PLoS One. 2020 Sep 11;15(9):e0238586. doi: 10.1371/journal.pone.0238586. eCollection 2020. PLoS One. 2020. PMID: 32915814 Free PMC article.
-
Computational modelling of the long-term effects of brain stimulation on the local and global structural connectivity of epileptic patients.PLoS One. 2020 Feb 6;15(2):e0221380. doi: 10.1371/journal.pone.0221380. eCollection 2020. PLoS One. 2020. PMID: 32027654 Free PMC article.
References
LinkOut - more resources
Full Text Sources
Other Literature Sources