

Mammalian target of rapamycin complex 1 signalling is essential for germinal centre reaction
- PMID: 28557002
- PMCID: PMC5588811
- DOI: 10.1111/imm.12767
Mammalian target of rapamycin complex 1 signalling is essential for germinal centre reaction
Abstract
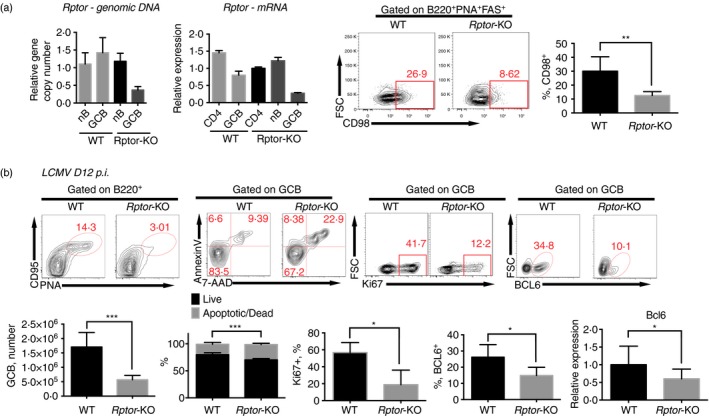
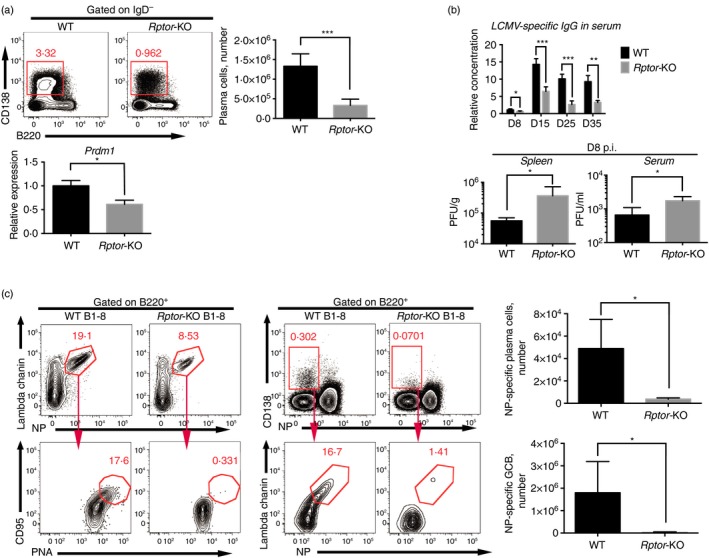
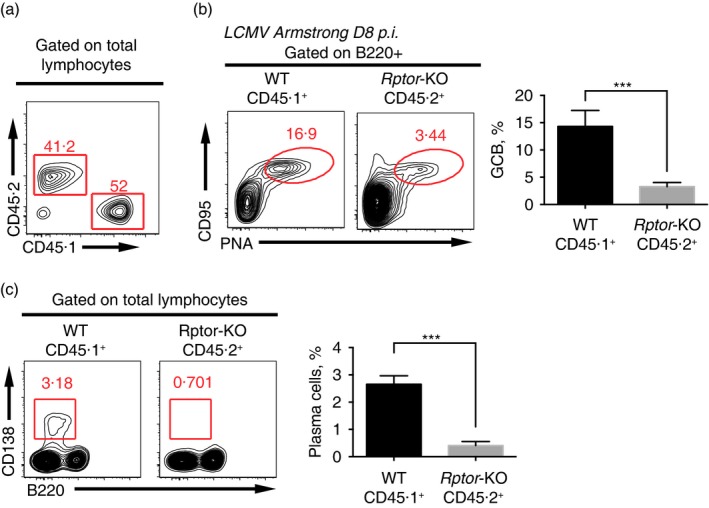
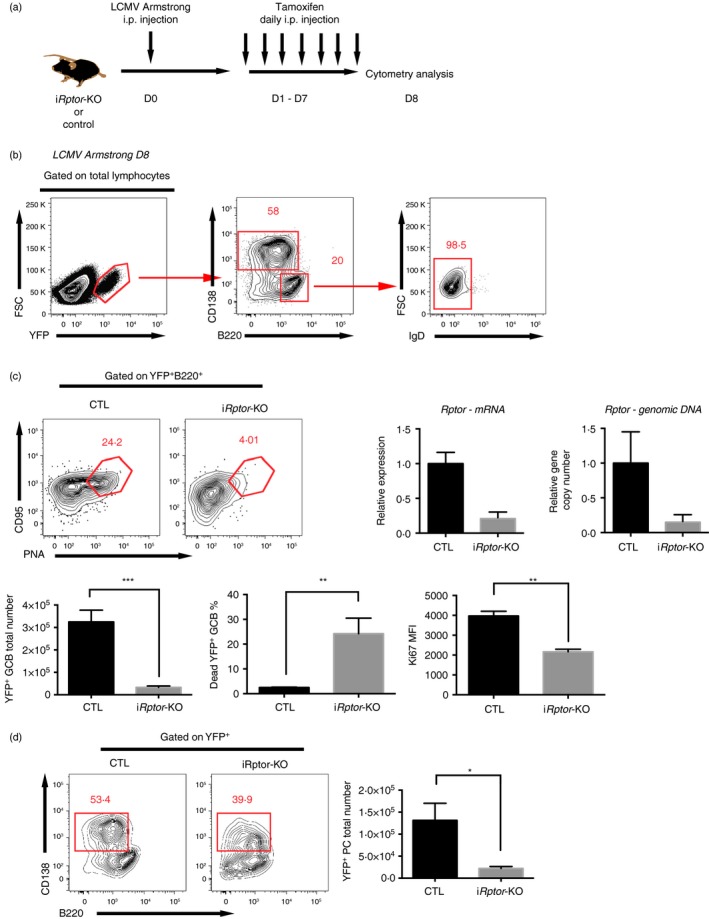
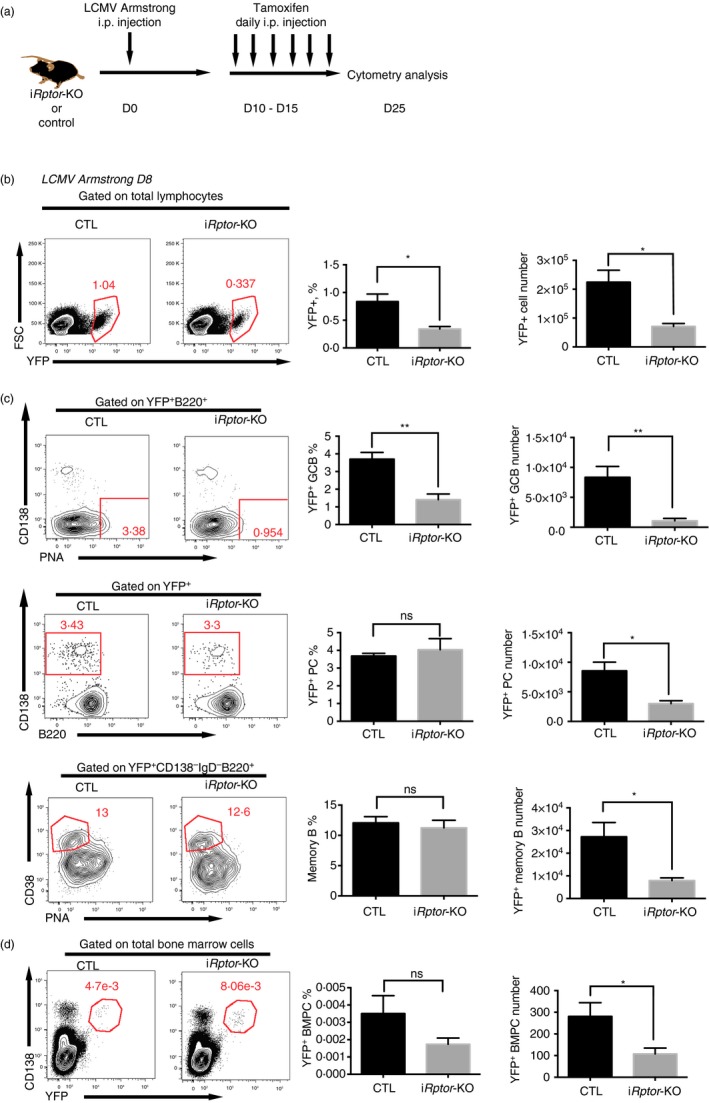
The mammalian target of rapamycin (mTOR) is a serine-threonine kinase that has been shown to be essential for the differentiation and function of various immune cells. Earlier in vitro studies showed that mTOR signalling regulates B-cell biology by supporting their activation and proliferation. However, how mTOR signalling temporally regulates in vivo germinal centre B (GCB) cell development and differentiation into short-lived plasma cells, long-lived plasma cells and memory cells is still not well understood. In this study, we used a combined conditional/inducible knock-out system to investigate the temporal regulation of mTOR complex 1 (mTORC1) in the GCB cell response to acute lymphocytic choriomeningitis virus infection by deleting Raptor, a main component of mTORC1, specifically in B cells in pre- and late GC phase. Early Raptor deficiency strongly inhibited GCB cell proliferation and differentiation and plasma cell differentiation. Nevertheless, late GC Raptor deficiency caused only decreases in the size of memory B cells and long-lived plasma cells through poor maintenance of GCB cells, but it did not change their differentiation. Collectively, our data revealed that mTORC1 signalling supports GCB cell responses at both early and late GC phases during viral infection but does not regulate GCB cell differentiation into memory B cells and plasma cells at the late GC stage.
Keywords: B cell; cell differentiation; gene regulation.
© 2017 John Wiley & Sons Ltd.
Figures
References
-
- Plotkin SA, Orenstein AW, Offit PA. Vaccines, 6th edn. Amsterdam, Netherlands: Elsevier Inc., 2012.
-
- Victora GD, Nussenzweig MC. Germinal centers. Annu Rev Immunol 2012; 30:429–57. - PubMed
-
- Neuberger MS, Lanoue A, Ehrenstein MR, Batista FD, Sale JE, Williams GT. Antibody diversification and selection in the mature B‐cell compartment. Cold Spring Harb Symp Quant Biol 1999; 64:211–6. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
