

SK4 channels modulate Ca2+ signalling and cell cycle progression in murine breast cancer
- PMID: 28557306
- PMCID: PMC5579333
- DOI: 10.1002/1878-0261.12087
SK4 channels modulate Ca2+ signalling and cell cycle progression in murine breast cancer
Abstract
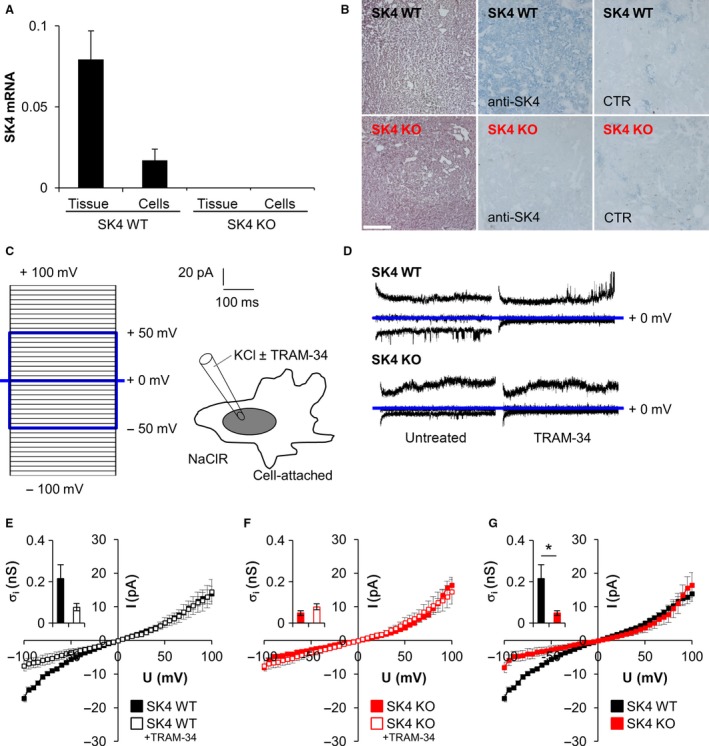
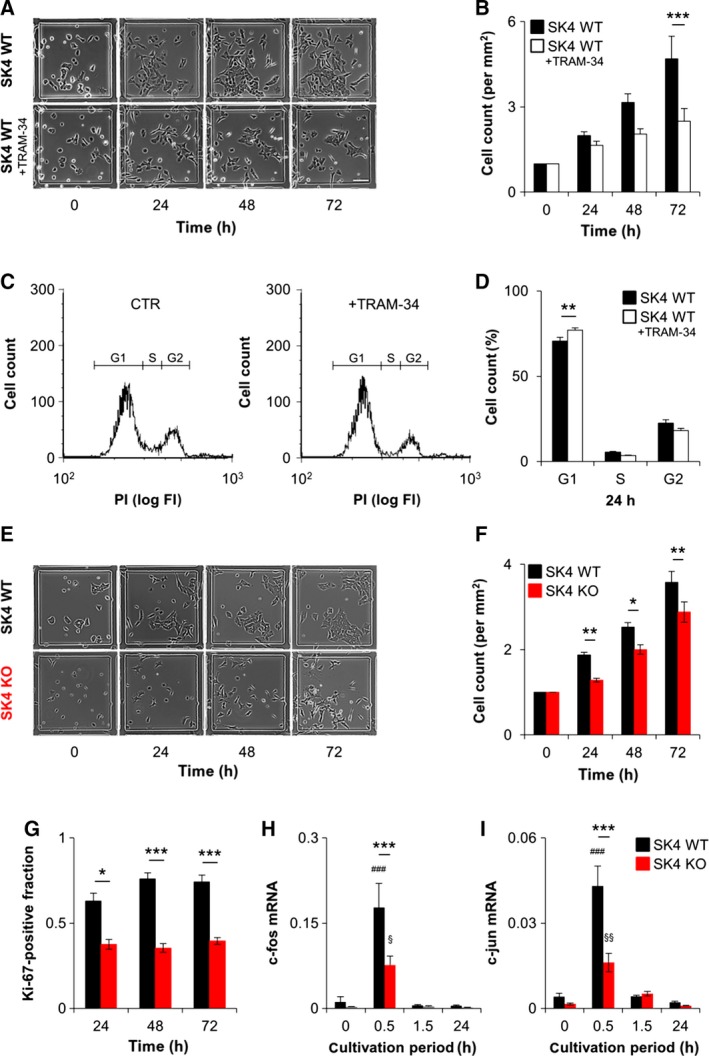
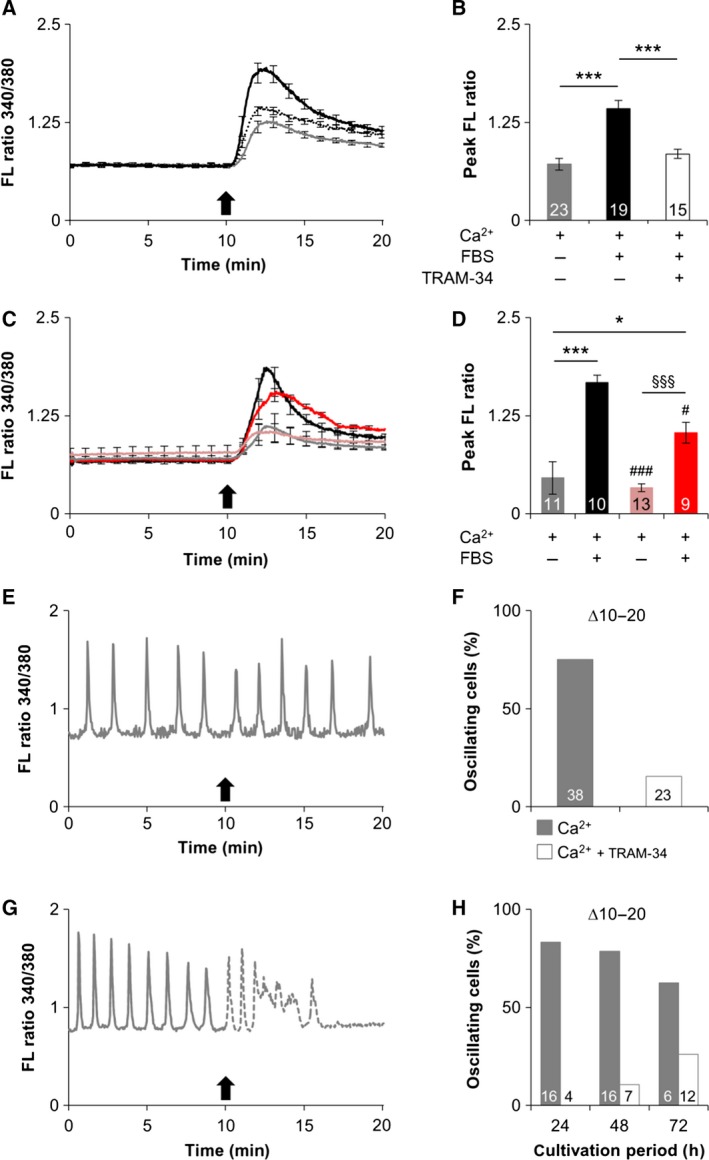
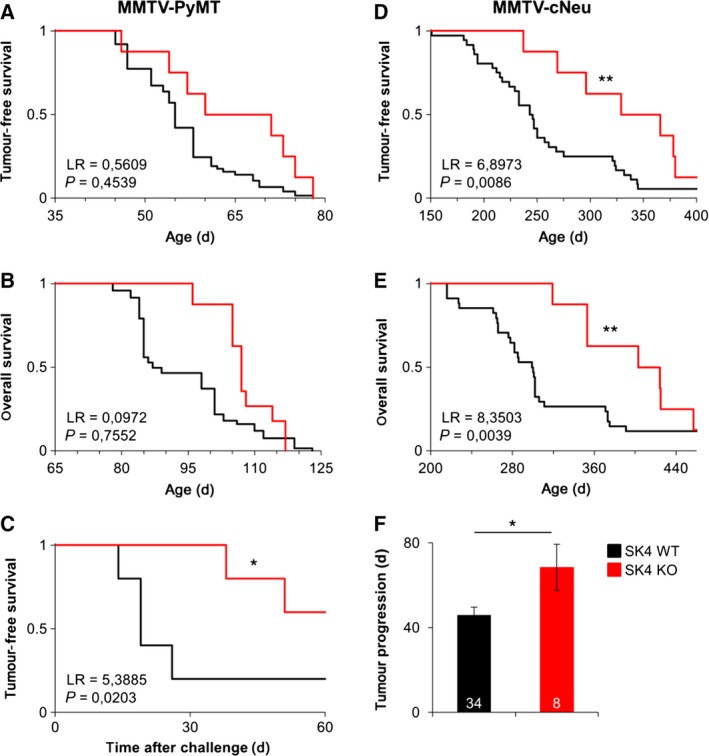
Oncogenic signalling via Ca2+ -activated K+ channels of intermediate conductance (SK4, also known as KCa 3.1 or IK) has been implicated in different cancer entities including breast cancer. Yet, the role of endogenous SK4 channels for tumorigenesis is unclear. Herein, we generated SK4-negative tumours by crossing SK4-deficient (SK4 KO) mice to the polyoma middle T-antigen (PyMT) and epidermal growth factor receptor 2 (cNeu) breast cancer models in which oncogene expression is driven by the retroviral promoter MMTV. Survival parameters and tumour progression were studied in cancer-prone SK4 KO in comparison with wild-type (WT) mice and in a syngeneic orthotopic mouse model following transplantation of SK4-negative or WT tumour cells. SK4 activity was modulated by genetic or pharmacological means using the SK4 inhibitor TRAM-34 in order to establish the role of breast tumour SK4 for cell growth, electrophysiological signalling, and [Ca2+ ]i oscillations. Ablation of SK4 and TRAM-34 treatment reduced the SK4-generated current fraction, growth factor-dependent Ca2+ entry, cell cycle progression and the proliferation rate of MMTV-PyMT tumour cells. In vivo, PyMT oncogene-driven tumorigenesis was only marginally affected by the global lack of SK4, whereas tumour progression was significantly delayed after orthotopic implantation of MMTV-PyMT SK4 KO breast tumour cells. However, overall survival and progression-free survival time in the MMTV-cNeu mouse model were significantly extended in the absence of SK4. Collectively, our data from murine breast cancer models indicate that SK4 activity is crucial for cell cycle control. Thus, the modulation of this channel should be further investigated towards a potential improvement of existing antitumour strategies in human breast cancer.
Keywords: Ca2+-activated K+ channels of intermediate conductance (SK4, KCa3.1, IK, KCNN4); breast cancer; epidermal growth factor receptor 2 (Her2/cNeu); mouse mammary tumour virus (MMTV); polyoma virus middle T-antigen (PyMT).
© 2017 The Authors. Published by FEBS Press and John Wiley & Sons Ltd.
Figures
Similar articles
-
KCa3.1 Channels and Glioblastoma: In Vitro Studies.Curr Neuropharmacol. 2018;16(5):627-635. doi: 10.2174/1570159X15666170808115821. Curr Neuropharmacol. 2018. PMID: 28786347 Free PMC article. Review.
-
KCa3.1 Channels Confer Radioresistance to Breast Cancer Cells.Cancers (Basel). 2019 Sep 1;11(9):1285. doi: 10.3390/cancers11091285. Cancers (Basel). 2019. PMID: 31480522 Free PMC article.
-
Subunits of BK channels promote breast cancer development and modulate responses to endocrine treatment in preclinical models.Br J Pharmacol. 2022 Jun;179(12):2906-2924. doi: 10.1111/bph.15147. Epub 2020 Aug 27. Br J Pharmacol. 2022. PMID: 32468618
-
Enhanced glucose tolerance by SK4 channel inhibition in pancreatic beta-cells.Diabetes. 2009 Aug;58(8):1835-43. doi: 10.2337/db08-1324. Epub 2009 Apr 28. Diabetes. 2009. PMID: 19401418 Free PMC article.
-
Intermediate conductance calcium-activated potassium channel (KCa3.1) in cancer: Emerging roles and therapeutic potentials.Biochem Pharmacol. 2024 Dec;230(Pt 1):116573. doi: 10.1016/j.bcp.2024.116573. Epub 2024 Oct 11. Biochem Pharmacol. 2024. PMID: 39396649 Review.
Cited by
-
Transcriptional Repression and Protein Degradation of the Ca2+-Activated K+ Channel KCa1.1 by Androgen Receptor Inhibition in Human Breast Cancer Cells.Front Physiol. 2018 Apr 16;9:312. doi: 10.3389/fphys.2018.00312. eCollection 2018. Front Physiol. 2018. PMID: 29713287 Free PMC article.
-
KCa3.1 Channels and Glioblastoma: In Vitro Studies.Curr Neuropharmacol. 2018;16(5):627-635. doi: 10.2174/1570159X15666170808115821. Curr Neuropharmacol. 2018. PMID: 28786347 Free PMC article. Review.
-
Cancer-Associated Intermediate Conductance Ca2+-Activated K⁺ Channel KCa3.1.Cancers (Basel). 2019 Jan 17;11(1):109. doi: 10.3390/cancers11010109. Cancers (Basel). 2019. PMID: 30658505 Free PMC article. Review.
-
Pathophysiological role of ion channels and transporters in HER2-positive breast cancer.Cancer Gene Ther. 2022 Aug;29(8-9):1097-1104. doi: 10.1038/s41417-021-00407-4. Epub 2022 Jan 7. Cancer Gene Ther. 2022. PMID: 34997219 Review.
-
KCa3.1 Channels Confer Radioresistance to Breast Cancer Cells.Cancers (Basel). 2019 Sep 1;11(9):1285. doi: 10.3390/cancers11091285. Cancers (Basel). 2019. PMID: 31480522 Free PMC article.
References
-
- Begenisich T, Nakamoto T, Ovitt CE, Nehrke K, Brugnara C, Alper SL and Melvin JE (2004) Physiological roles of the intermediate conductance, Ca2 + ‐activated potassium channel Kcnn4. J Biol Chem 279, 47681–47687. - PubMed
-
- Bulk E, Ay AS, Hammadi M, Ouadid‐Ahidouch H, Schelhaas S, Hascher A, Rohde C, Thoennissen NH, Wiewrodt R, Schmidt E et al (2015) Epigenetic dysregulation of KCa 3.1 channels induces poor prognosis in lung cancer. Int J Cancer 137, 1306–1317. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
