

Bacteria exploit a polymorphic instability of the flagellar filament to escape from traps
- PMID: 28559324
- PMCID: PMC5474801
- DOI: 10.1073/pnas.1701644114
Bacteria exploit a polymorphic instability of the flagellar filament to escape from traps
Abstract
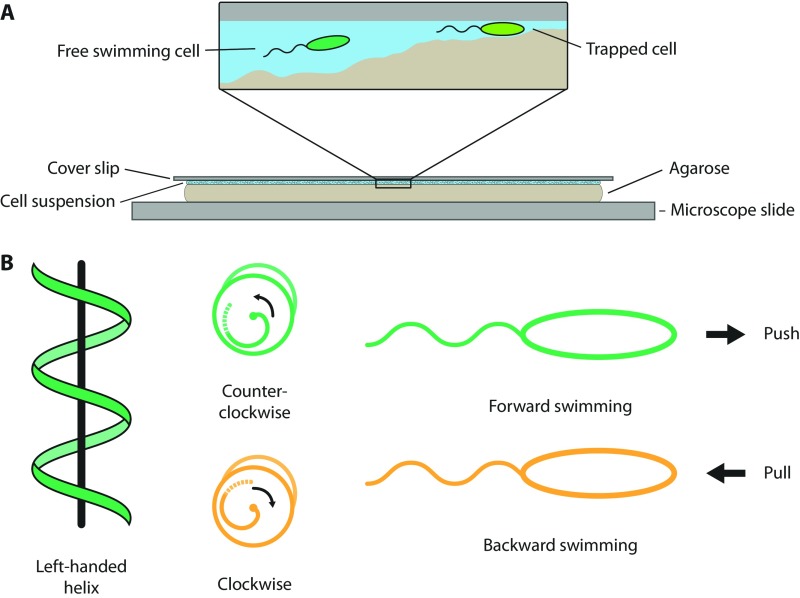
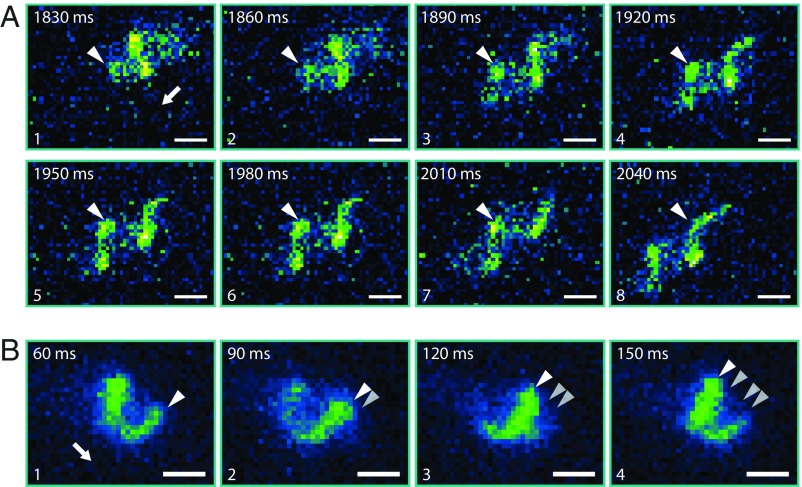
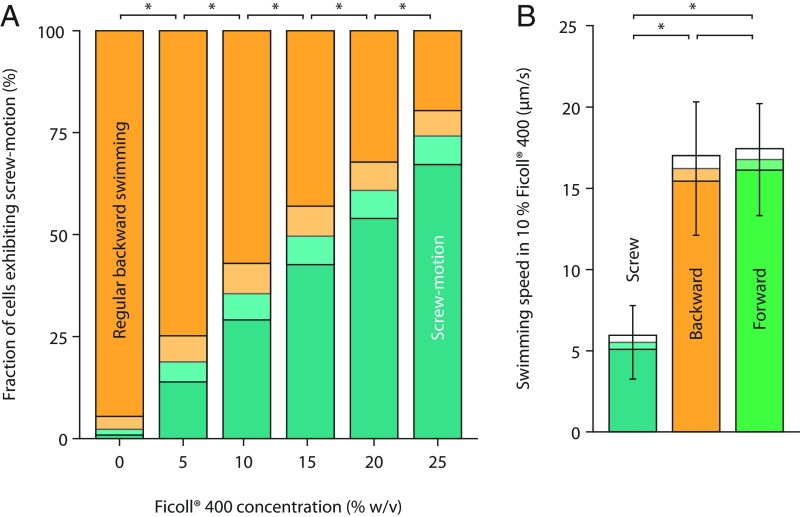
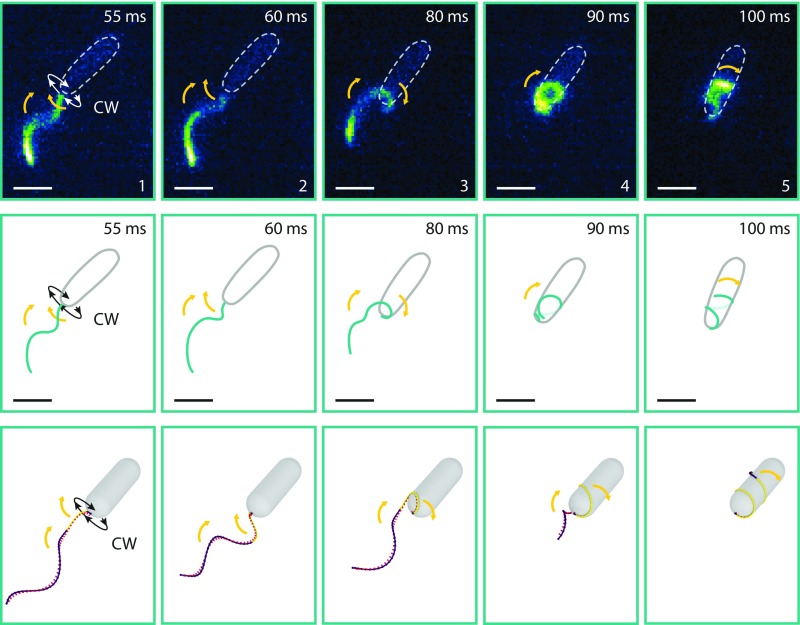
Many bacterial species swim by rotating single polar helical flagella. Depending on the direction of rotation, they can swim forward or backward and change directions to move along chemical gradients but also to navigate their obstructed natural environment in soils, sediments, or mucus. When they get stuck, they naturally try to back out, but they can also resort to a radically different flagellar mode, which we discovered here. Using high-speed microscopy, we monitored the swimming behavior of the monopolarly flagellated species Shewanella putrefaciens with fluorescently labeled flagellar filaments at an agarose-glass interface. We show that, when a cell gets stuck, the polar flagellar filament executes a polymorphic change into a spiral-like form that wraps around the cell body in a spiral-like fashion and enables the cell to escape by a screw-like backward motion. Microscopy and modeling suggest that this propagation mode is triggered by an instability of the flagellum under reversal of the rotation and the applied torque. The switch is reversible and bacteria that have escaped the trap can return to their normal swimming mode by another reversal of motor direction. The screw-type flagellar arrangement enables a unique mode of propagation and, given the large number of polarly flagellated bacteria, we expect it to be a common and widespread escape or motility mode in complex and structured environments.
Keywords: Shewanella; flagella; motility; structured environment.
Conflict of interest statement
The authors declare no conflict of interest.
Figures



References
-
- Kelly FX, Dapsis KJ, Lauffenburger DA. Effect of bacterial chemotaxis on dynamics of microbial competition. Microb Ecol. 1988;16:115–131. - PubMed
-
- Lauffenburger DA. Quantitative studies of bacterial chemotaxis and microbial population dynamics. Microb Ecol. 1991;22:175–185. - PubMed
-
- Fenchel T. Microbial behavior in a heterogeneous world. Science. 2002;296:1068–1071. - PubMed
-
- Alexandre G, Greer-Phillips S, Zhulin IB. Ecological role of energy taxis in microorganisms. FEMS Microbiol Rev. 2004;28:113–126. - PubMed
-
- Reichenbach T, Mobilia M, Frey E. Mobility promotes and jeopardizes biodiversity in rock-paper-scissors games. Nature. 2007;448:1046–1049. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
