

Input timing for spatial processing is precisely tuned via constant synaptic delays and myelination patterns in the auditory brainstem
- PMID: 28559325
- PMCID: PMC5474802
- DOI: 10.1073/pnas.1702290114
Input timing for spatial processing is precisely tuned via constant synaptic delays and myelination patterns in the auditory brainstem
Abstract
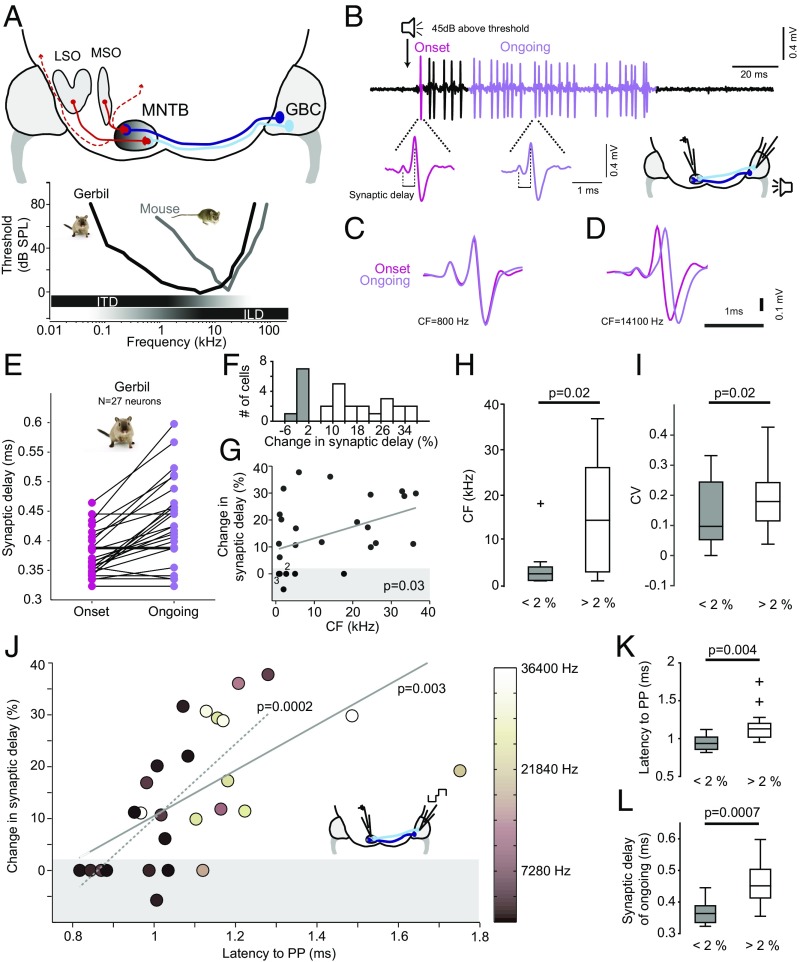
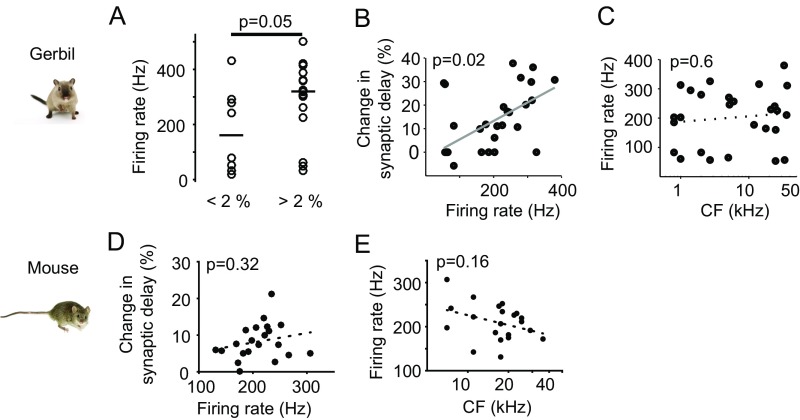
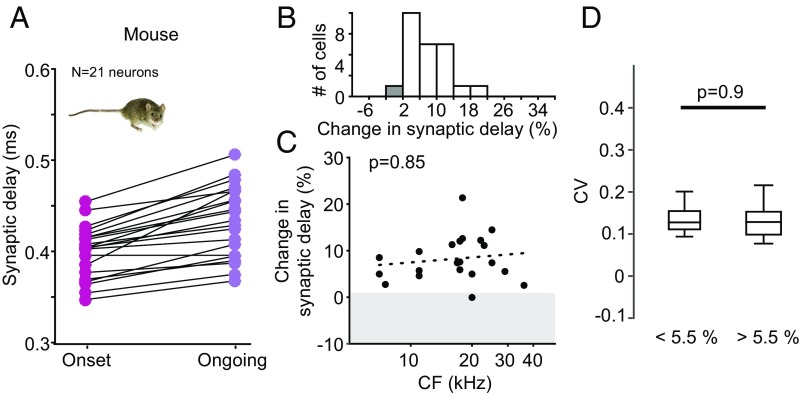
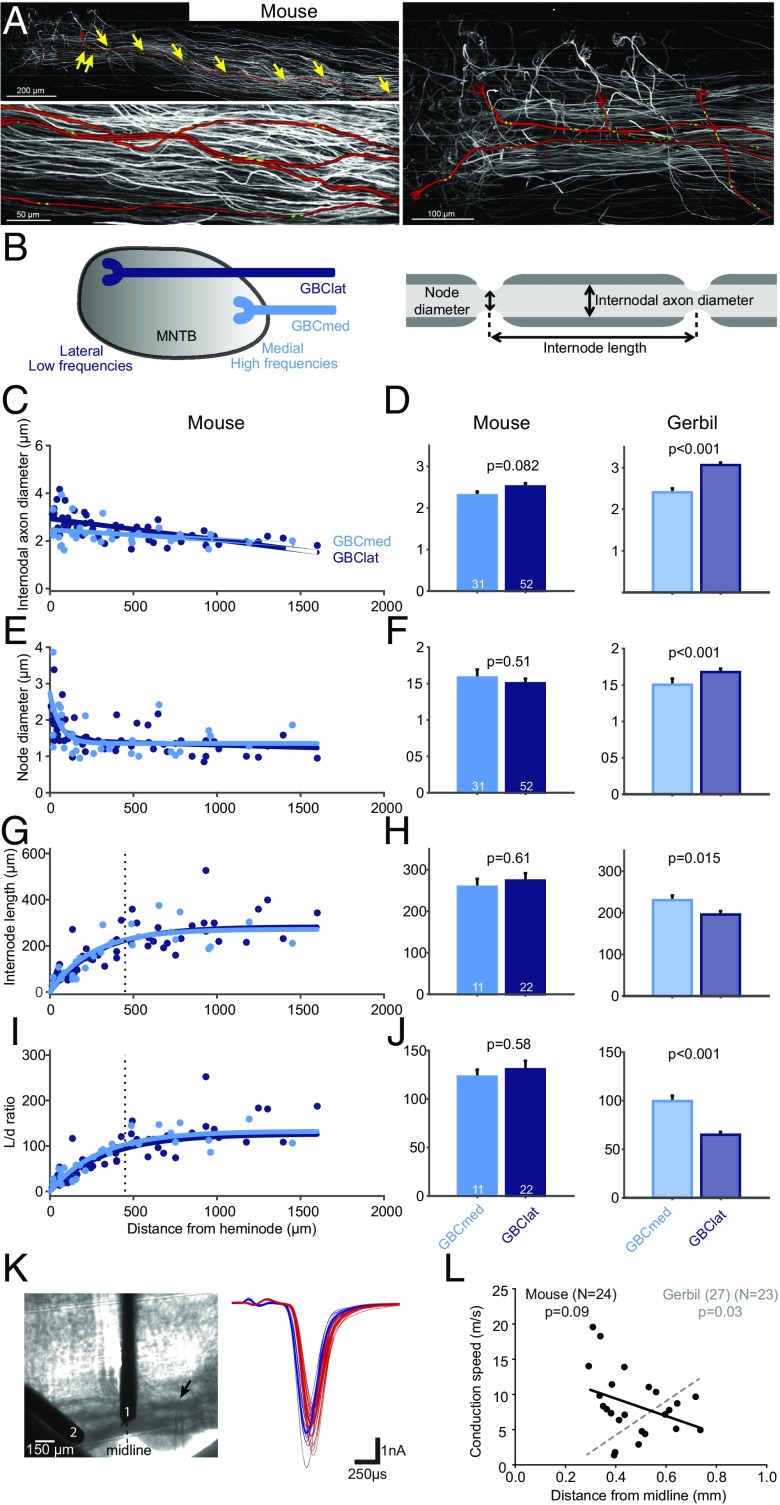
Precise timing of synaptic inputs is a fundamental principle of neural circuit processing. The temporal precision of postsynaptic input integration is known to vary with the computational requirements of a circuit, yet how the timing of action potentials is tuned presynaptically to match these processing demands is not well understood. In particular, action potential timing is shaped by the axonal conduction velocity and the duration of synaptic transmission delays within a pathway. However, it is not known to what extent these factors are adapted to the functional constraints of the respective circuit. Here, we report the finding of activity-invariant synaptic transmission delays as a functional adaptation for input timing adjustment in a brainstem sound localization circuit. We compared axonal and synaptic properties of the same pathway between two species with dissimilar timing requirements (gerbil and mouse): In gerbils (like humans), neuronal processing of sound source location requires exceptionally high input precision in the range of microseconds, but not in mice. Activity-invariant synaptic transmission and conduction delays were present exclusively in fast conducting axons of gerbils that also exhibited unusual structural adaptations in axon myelination for increased conduction velocity. In contrast, synaptic transmission delays in mice varied depending on activity levels, and axonal myelination and conduction velocity exhibited no adaptations. Thus, the specializations in gerbils and their absence in mice suggest an optimization of axonal and synaptic properties to the specific demands of sound localization. These findings significantly advance our understanding of structural and functional adaptations for circuit processing.
Keywords: circuit processing; input timing; myelination; sound localization; synaptic transmission delay.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Wehr M, Zador AM. Balanced inhibition underlies tuning and sharpens spike timing in auditory cortex. Nature. 2003;426:442–446. - PubMed
-
- Pouille F, Scanziani M. Enforcement of temporal fidelity in pyramidal cells by somatic feed-forward inhibition. Science. 2001;293:1159–1163. - PubMed
-
- Brand A, Behrend O, Marquardt T, McAlpine D, Grothe B. Precise inhibition is essential for microsecond interaural time difference coding. Nature. 2002;417:543–547. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
