

Finding the Needle in the Haystack-the Use of Microfluidic Droplet Technology to Identify Vitamin-Secreting Lactic Acid Bacteria
- PMID: 28559484
- PMCID: PMC5449655
- DOI: 10.1128/mBio.00526-17
Finding the Needle in the Haystack-the Use of Microfluidic Droplet Technology to Identify Vitamin-Secreting Lactic Acid Bacteria
Abstract
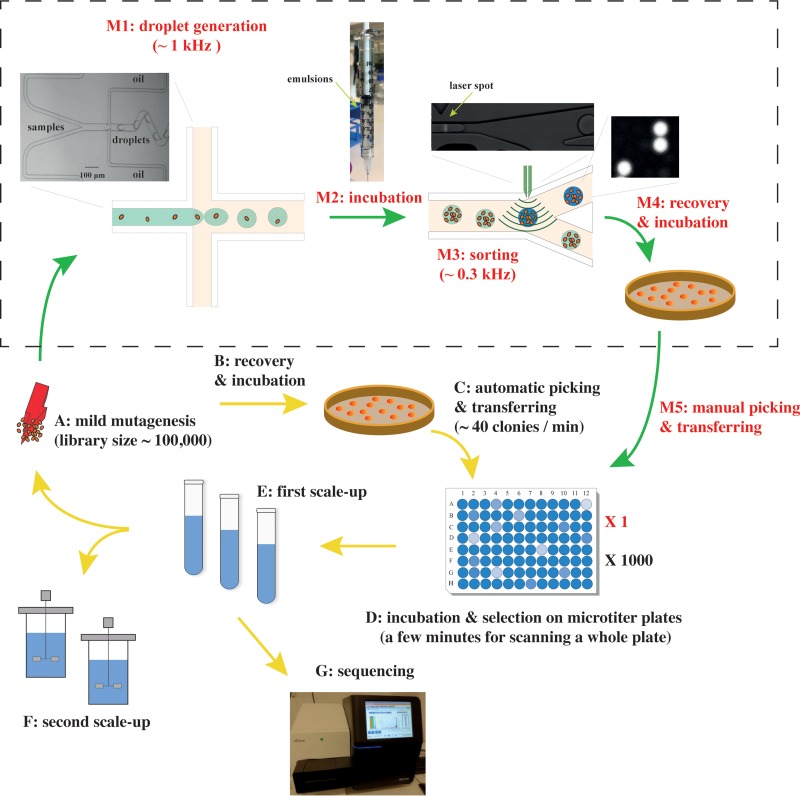
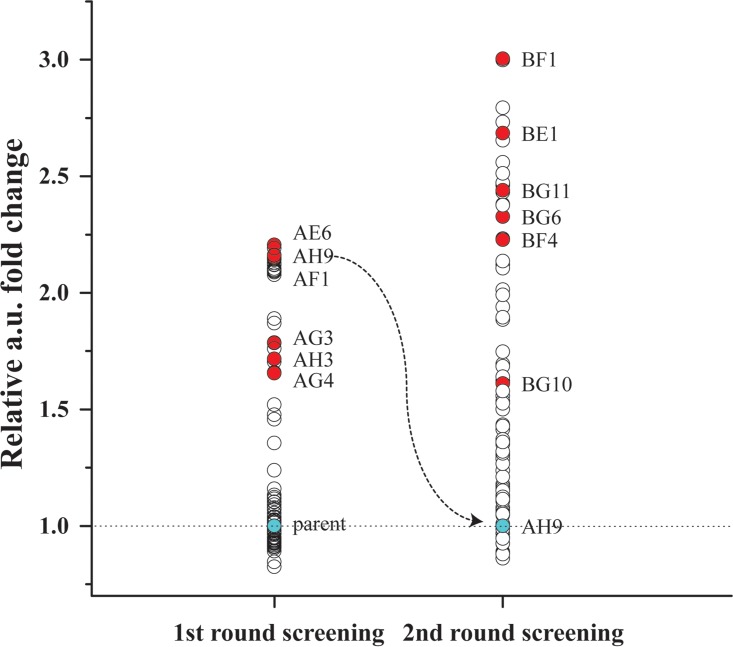
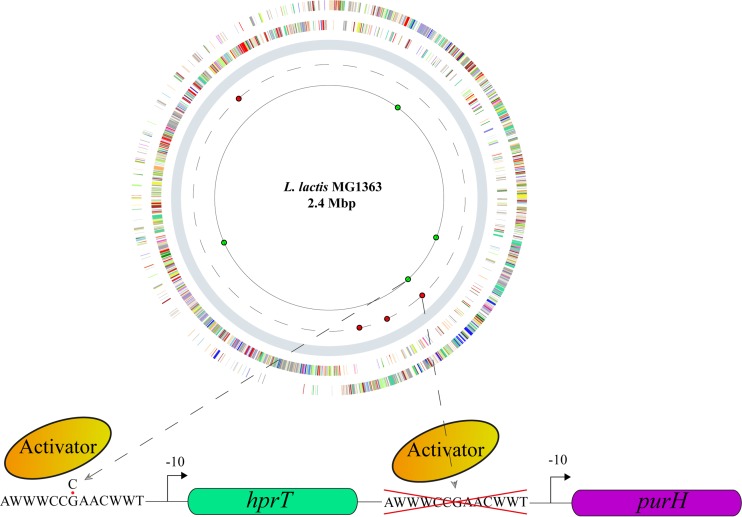
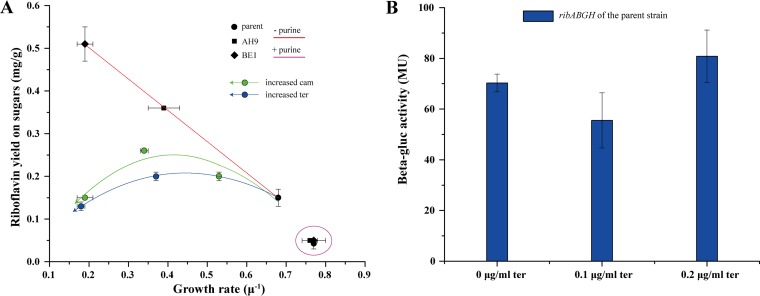
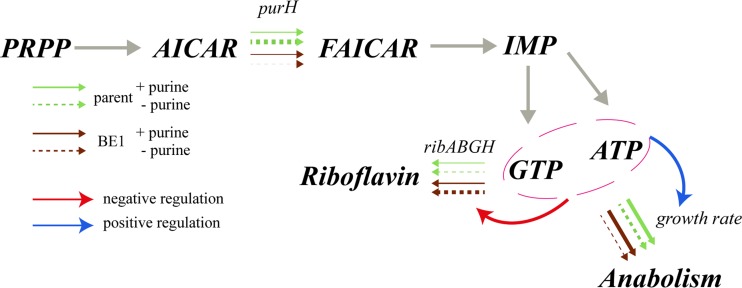
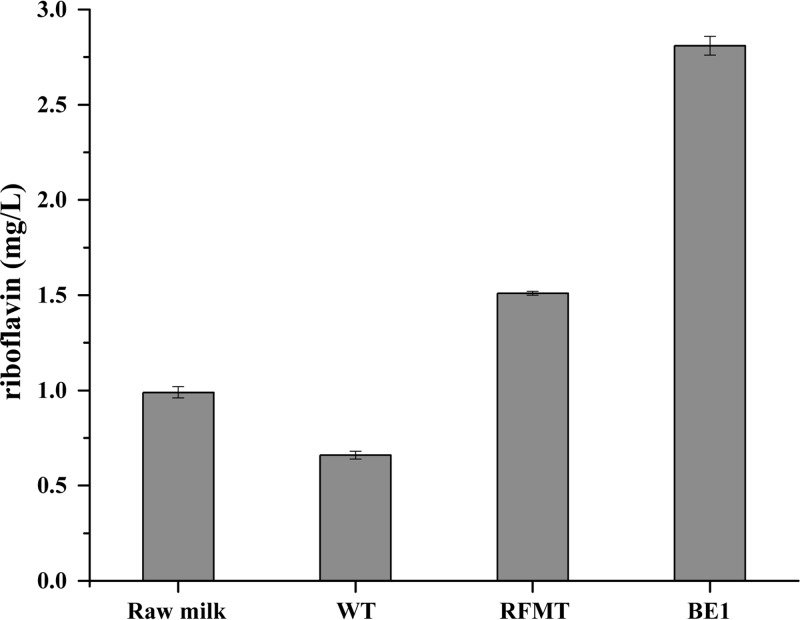
Efficient screening technologies aim to reduce both the time and the cost required for identifying rare mutants possessing a phenotype of interest in a mutagenized population. In this study, we combined a mild mutagenesis strategy with high-throughput screening based on microfluidic droplet technology to identify Lactococcus lactis variants secreting vitamin B2 (riboflavin). Initially, we used a roseoflavin-resistant mutant of L. lactis strain MG1363, JC017, which secreted low levels of riboflavin. By using fluorescence-activated droplet sorting, several mutants that secreted riboflavin more efficiently than JC017 were readily isolated from the mutagenesis library. The screening was highly efficient, and candidates with as few as 1.6 mutations per million base pairs (Mbp) were isolated. The genetic characterization revealed that riboflavin production was triggered by mutations inhibiting purine biosynthesis, which is surprising since the purine nucleotide GTP is a riboflavin precursor. Purine starvation in the mutants induced overexpression of the riboflavin biosynthesis cluster ribABGH When the purine starvation was relieved by purine supplementation in the growth medium, the outcome was an immediate downregulation of the riboflavin biosynthesis cluster and a reduction in riboflavin production. Finally, by applying the new isolates in milk fermentation, the riboflavin content of milk (0.99 mg/liter) was improved to 2.81 mg/liter, compared with 0.66 mg/liter and 1.51 mg/liter by using the wild-type strain and the original roseoflavin-resistant mutant JC017, respectively. The results obtained demonstrate how powerful classical mutagenesis can be when combined with droplet-based microfluidic screening technology for obtaining microorganisms with useful attributes.IMPORTANCE The food industry prefers to use classical approaches, e.g., random mutagenesis followed by screening, to improve microorganisms used in food production, as the use of recombinant DNA technologies is still not widely accepted. Although modern automated screening platforms are widely accessible, screening remains as a bottleneck in strain development, especially when a mild mutagenesis approach is applied to reduce the chance of accumulating unintended mutations, which may cause unwanted phenotypic changes. Here, we incorporate a droplet-based high-throughput screening method into the strain development process and readily capture L. lactis variants with more efficient vitamin secretion from low-error-rate mutagenesis libraries. This study shows that useful mutants showing strong phenotypes but without extensive mutations can be identified with efficient screening technologies. It is therefore possible to avoid accumulating detrimental mutations while enriching beneficial ones through iterative mutagenesis screening. Due to the low mutation rates, the genetic determinants are also readily identified.
Keywords: classical mutagenesis; droplet microfluidics; high-throughput screening; lactic acid bacteria; vitamin B2.
Copyright © 2017 Chen et al.
Figures
References
-
- Stephanopoulos G, Aristidou AA, Nielsen J. 1998. Metabolic engineering: principles and methodologies. Academic Press, New York, NY.
-
- Johansen E, Øregaard G, Sørensen KI, Derkx PMF. 2015. Modern approaches for isolation, selection, and improvement of bacterial strains for fermentation applications, p 227–248. In Holzapfel W (ed), Advances in fermented foods and beverages: improving quality, technologies and health benefits. A volume in Woodhead Publishing Series in Food Science, Technology and Nutrition. Woodhead Publishing, Cambridge, United Kingdom. doi: 10.1016/B978-1-78242-015-6.00010-4. - DOI
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
