

Threshold position control of anticipation in humans: a possible role of corticospinal influences
- PMID: 28560812
- PMCID: PMC5538209
- DOI: 10.1113/JP274309
Threshold position control of anticipation in humans: a possible role of corticospinal influences
Abstract
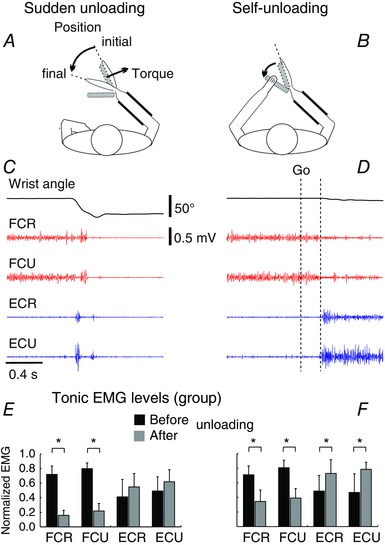
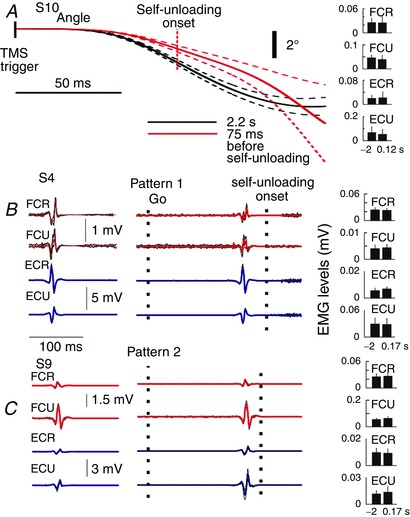
Key points: Sudden unloading of preloaded wrist muscles elicits motion to a new wrist position. Such motion is prevented if subjects unload muscles using the contralateral arm (self-unloading). Corticospinal influences originated from the primary motor cortex maintain tonic influences on motoneurons of wrist muscles before sudden unloading but modify these influences prior to the onset and until the end of self-unloading. Results are interpreted based on the previous finding that intentional actions are caused by central, particularly corticospinal, shifts in the spatial thresholds at which wrist motoneurons are activated, thus predetermining the attractor point at which the neuromuscular periphery achieves mechanical balance with environment forces. By maintaining or shifting the thresholds, descending systems let body segments go to the equilibrium position in the respective unloading tasks without the pre-programming of kinematics or muscle activation patterns. The study advances the understanding of how motor actions in general, and anticipation in particular, are controlled.
Abstract: The role of corticospinal (CS) pathways in anticipatory motor actions was evaluated using transcranial magnetic stimulation (TMS) of the primary motor cortex projecting to motoneurons (MNs) of wrist muscles. Preloaded wrist flexors were suddenly unloaded by the experimenter or by the subject using the other hand (self-unloading). After sudden unloading, the wrist joint involuntarily flexed to a new position. In contrast, during self-unloading the wrist remained almost motionless, implying that an anticipatory postural adjustment occurred. In the self-unloading task, anticipation was manifested by a decrease in descending facilitation of pre-activated flexor MNs starting ∼72 ms before changes in the background EMG activity. Descending facilitation of extensor MNs began to increase ∼61 ms later. Conversely, these influences remained unchanged before sudden unloading, implying the absence of anticipation. We also tested TMS responses during EMG silent periods produced by brief muscle shortening, transiently resulting in similar EMG levels before the onset and after the end of self-unloading. We found reduced descending facilitation of flexor MNs after self-unloading. To explain why the wrist excursion was minimized in self-unloading due to these changes in descending influences, we relied on previous demonstrations that descending systems pre-set the threshold positions of body segments at which muscles begin to be activated, thus predetermining the equilibrium point to which the system is attracted. Based on this notion, a more consistent explanation of the kinematic, EMG and descending patterns in the two types of unloading is proposed compared to the alternative notion of direct pre-programming of kinematic and/or EMG patterns.
Keywords: behavioural neuroscience; control variables; motor cortex.
© 2017 Université de Montréal. The Journal of Physiology © 2017 The Physiological Society.
Figures





Similar articles
-
Subthreshold corticospinal control of anticipatory actions in humans.Behav Brain Res. 2011 Oct 10;224(1):145-54. doi: 10.1016/j.bbr.2011.05.041. Epub 2011 Jun 7. Behav Brain Res. 2011. PMID: 21672559
-
Control of wrist position and muscle relaxation by shifting spatial frames of reference for motoneuronal recruitment: possible involvement of corticospinal pathways.J Physiol. 2010 May 1;588(Pt 9):1551-70. doi: 10.1113/jphysiol.2009.186858. Epub 2010 Mar 15. J Physiol. 2010. PMID: 20231141 Free PMC article.
-
Corticospinal control strategies underlying voluntary and involuntary wrist movements.Behav Brain Res. 2013 Jan 1;236(1):350-358. doi: 10.1016/j.bbr.2012.09.008. Epub 2012 Sep 13. Behav Brain Res. 2013. PMID: 22983216
-
Modulation of corticospinal influence over hand muscles during gripping tasks in man and monkey.Can J Physiol Pharmacol. 1996 Apr;74(4):547-58. Can J Physiol Pharmacol. 1996. PMID: 8828899 Review.
-
Contributions to the understanding of gait control.Dan Med J. 2014 Apr;61(4):B4823. Dan Med J. 2014. PMID: 24814597 Review.
Cited by
-
Indirect, referent control of motor actions underlies directional tuning of neurons.J Neurophysiol. 2019 Mar 1;121(3):823-841. doi: 10.1152/jn.00575.2018. Epub 2018 Dec 19. J Neurophysiol. 2019. PMID: 30565957 Free PMC article. Review.
-
Eye and head movements and vestibulo-ocular reflex in the context of indirect, referent control of motor actions.J Neurophysiol. 2020 Jul 1;124(1):115-133. doi: 10.1152/jn.00076.2020. Epub 2020 Jun 3. J Neurophysiol. 2020. PMID: 32490708 Free PMC article. Review.
-
Vestibular and corticospinal control of human body orientation in the gravitational field.J Neurophysiol. 2018 Dec 1;120(6):3026-3041. doi: 10.1152/jn.00483.2018. Epub 2018 Sep 12. J Neurophysiol. 2018. PMID: 30207862 Free PMC article.
-
Participation of ipsilateral cortical descending influences in bimanual wrist movements in humans.Exp Brain Res. 2020 Oct;238(10):2359-2372. doi: 10.1007/s00221-020-05899-4. Epub 2020 Aug 6. Exp Brain Res. 2020. PMID: 32766959
-
Referent control of the orientation of posture and movement in the gravitational field.Exp Brain Res. 2018 Feb;236(2):381-398. doi: 10.1007/s00221-017-5133-y. Epub 2017 Nov 21. Exp Brain Res. 2018. PMID: 29164285
References
-
- Asatryan DG & Feldman AG (1965). Functional tuning of the nervous system with control of movement or maintenance of a steady posture – I. Mechanographic analysis of the work of the joint during the performance of a postural task. Biophysics 10, 925–935.
-
- Belenkii V, Gurfinkel VS & Palstev YI (1967). On the elements of control of voluntary movements. Biophysics 12, 154–160.
-
- Bestmann S & Krakauer JW (2015). The uses and interpretations of the motor‐evoked potential for understanding behaviour. Exp Brain Res 233, 679–689. - PubMed
-
- Capaday C, Devanne H, Bertrand L & Lavoie BA (1998). Intracortical connections between motor cortical zones controlling antagonistic muscles in the cat: a combined anatomical and physiological study. Exp Brain Res 120, 223–232. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources