

Impaired activity of adherens junctions contributes to endothelial dilator dysfunction in ageing rat arteries
- PMID: 28561330
- PMCID: PMC5538197
- DOI: 10.1113/JP274189
Impaired activity of adherens junctions contributes to endothelial dilator dysfunction in ageing rat arteries
Abstract
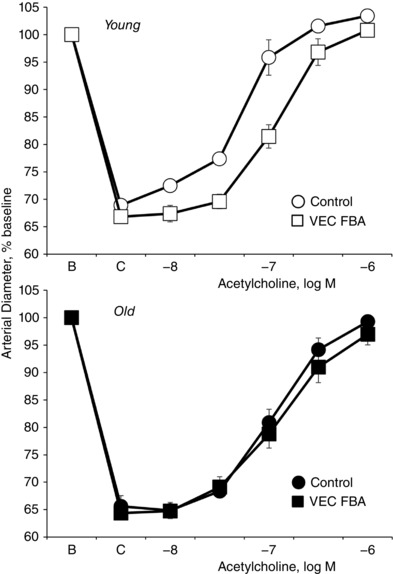
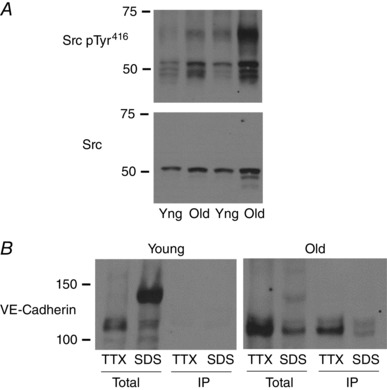
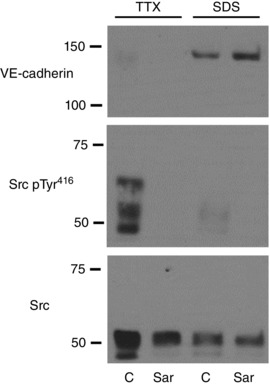
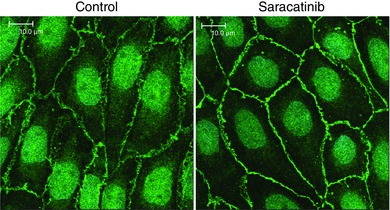
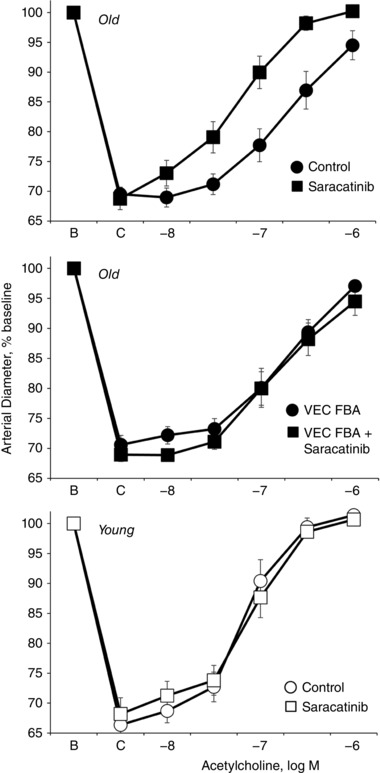
Key points: Ageing-induced endothelial dysfunction contributes to organ dysfunction and progression of cardiovascular disease. VE-cadherin clustering at adherens junctions promotes protective endothelial functions, including endothelium-dependent dilatation. Ageing increased internalization and degradation of VE-cadherin, resulting in impaired activity of adherens junctions. Inhibition of VE-cadherin clustering at adherens junctions (function-blocking antibody; FBA) reduced endothelial dilatation in young arteries but did not affect the already impaired dilatation in old arteries. After junctional disruption with the FBA, dilatation was similar in young and old arteries. Src tyrosine kinase activity and tyrosine phosphorylation of VE-cadherin were increased in old arteries. Src inhibition increased VE-cadherin at adherens junctions and increased endothelial dilatation in old, but not young, arteries. Src inhibition did not increase dilatation in old arteries treated with the VE-cadherin FBA. Ageing impairs the activity of adherens junctions, which contributes to endothelial dilator dysfunction. Restoring the activity of adherens junctions could be of therapeutic benefit in vascular ageing.
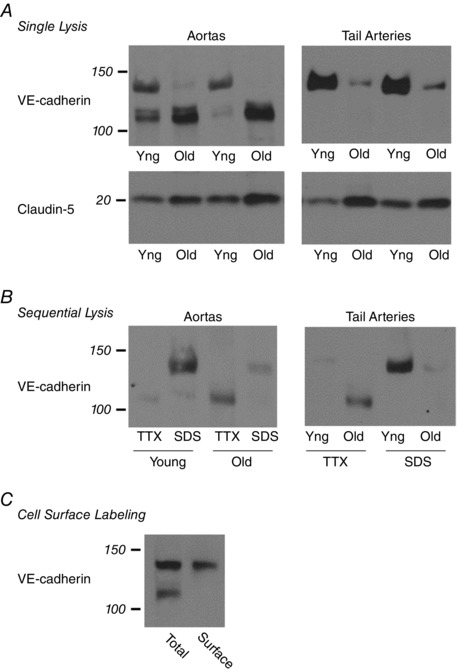
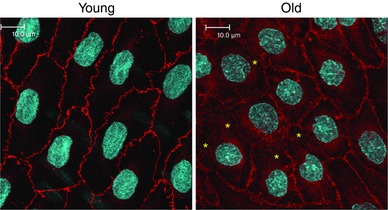
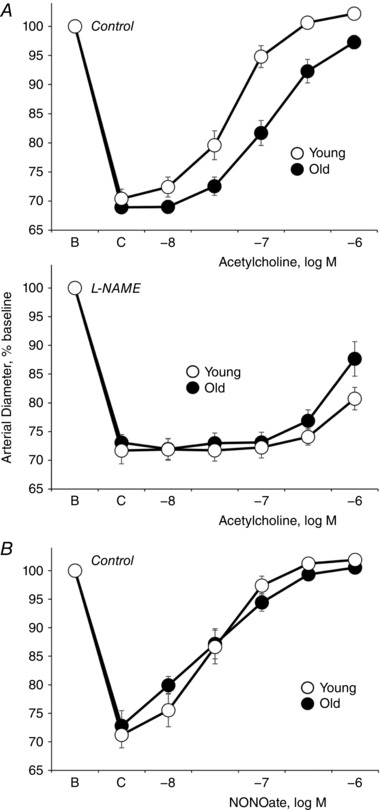
Abstract: Endothelial dilator dysfunction contributes to pathological vascular ageing. Experiments assessed whether altered activity of endothelial adherens junctions (AJs) might contribute to this dysfunction. Aortas and tail arteries were isolated from young (3-4 months) and old (22-24 months) F344 rats. VE-cadherin immunofluorescent staining at endothelial AJs and AJ width were reduced in old compared to young arteries. A 140 kDa VE-cadherin species was present on the cell surface and in TTX-insoluble fractions, consistent with junctional localization. Levels of the 140 kDa VE-cadherin were decreased, whereas levels of a TTX-soluble 115 kDa VE-cadherin species were increased in old compared to young arteries. Acetylcholine caused endothelium-dependent dilatation that was decreased in old compared to young arteries. Disruption of VE-cadherin clustering at AJs (function-blocking antibody, FBA) inhibited dilatation to acetylcholine in young, but not old, arteries. After the FBA, there was no longer any difference in dilatation between old and young arteries. Src activity and tyrosine phosphorylation of VE-cadherin were increased in old compared to young arteries. In old arteries, Src inhibition (saracatinib) increased: (i) 140 kDa VE-cadherin in the TTX-insoluble fraction, (ii) VE-cadherin intensity at AJs, (iii) AJ width, and (iv) acetylcholine dilatation. In old arteries treated with the FBA, saracatinib no longer increased acetylcholine dilatation. Saracatinib did not affect dilatation in young arteries. Therefore, ageing impairs AJ activity, which appears to reflect Src-induced phosphorylation, internalization and degradation of VE-cadherin. Moreover, impaired AJ activity can account for the endothelial dilator dysfunction in old arteries. Restoring endothelial AJ activity may be a novel therapeutic approach to vascular ageing.
Keywords: NO; VE-cadherin; adherens junctions; aging; endothelium.
© 2017 The Authors. The Journal of Physiology © 2017 The Physiological Society.
Figures
Comment in
-
Ageing-induced internalization and degradation of VE-cadherin leads to vascular dysfunction.J Physiol. 2018 Jan 15;596(2):143-144. doi: 10.1113/JP275495. Epub 2017 Dec 28. J Physiol. 2018. PMID: 29210069 Free PMC article. No abstract available.
Similar articles
-
Superoxide inhibition restores endothelium-dependent dilatation in aging arteries by enhancing impaired adherens junctions.Am J Physiol Heart Circ Physiol. 2018 Apr 1;314(4):H805-H811. doi: 10.1152/ajpheart.00681.2017. Epub 2018 Jan 12. Am J Physiol Heart Circ Physiol. 2018. PMID: 29351453 Free PMC article.
-
The role of adherens junctions and VE-cadherin in the control of vascular permeability.J Cell Sci. 2008 Jul 1;121(Pt 13):2115-22. doi: 10.1242/jcs.017897. J Cell Sci. 2008. PMID: 18565824 Review.
-
Pressure-induced maturation of endothelial cells on newborn mouse carotid arteries.Am J Physiol Heart Circ Physiol. 2013 Aug 1;305(3):H321-9. doi: 10.1152/ajpheart.00099.2013. Epub 2013 May 24. Am J Physiol Heart Circ Physiol. 2013. PMID: 23709593 Free PMC article.
-
RAB26-dependent autophagy protects adherens junctional integrity in acute lung injury.Autophagy. 2018;14(10):1677-1692. doi: 10.1080/15548627.2018.1476811. Epub 2018 Jul 26. Autophagy. 2018. PMID: 29965781 Free PMC article.
-
Dynamic Regulation of Vascular Permeability by Vascular Endothelial Cadherin-Mediated Endothelial Cell-Cell Junctions.J Nippon Med Sch. 2017;84(4):148-159. doi: 10.1272/jnms.84.148. J Nippon Med Sch. 2017. PMID: 28978894 Review.
Cited by
-
Evaluation of Serum Endocan Levels in Sensorineural Hearing Loss.J Int Adv Otol. 2019 Apr;15(1):83-86. doi: 10.5152/iao.2018.5051. J Int Adv Otol. 2019. PMID: 30325334 Free PMC article.
-
Reactive species-induced microvascular dysfunction in ischemia/reperfusion.Free Radic Biol Med. 2019 May 1;135:182-197. doi: 10.1016/j.freeradbiomed.2019.02.031. Epub 2019 Mar 5. Free Radic Biol Med. 2019. PMID: 30849489 Free PMC article. Review.
-
Dynamic Crosstalk between Vascular Smooth Muscle Cells and the Aged Extracellular Matrix.Int J Mol Sci. 2021 Sep 21;22(18):10175. doi: 10.3390/ijms221810175. Int J Mol Sci. 2021. PMID: 34576337 Free PMC article. Review.
-
Aging modulates homeostatic leukocyte trafficking to the peritoneal cavity in a sex-specific manner.J Leukoc Biol. 2023 Sep 27;114(4):301-314. doi: 10.1093/jleuko/qiad053. J Leukoc Biol. 2023. PMID: 37309034 Free PMC article.
-
Evolving structure-function relations during aortic maturation and aging revealed by multiphoton microscopy.Mech Ageing Dev. 2021 Jun;196:111471. doi: 10.1016/j.mad.2021.111471. Epub 2021 Mar 16. Mech Ageing Dev. 2021. PMID: 33741396 Free PMC article.
References
-
- Angelini DJ, Hyun SW, Grigoryev DN, Garg P, Gong P, Singh IS, Passaniti A, Hasday JD & Goldblum SE (2006). TNF‐α increases tyrosine phosphorylation of vascular endothelial cadherin and opens the paracellular pathway through fyn activation in human lung endothelia. Am J Physiol Lung Cell Mol Physiol 291, L1232–L1245. - PubMed
-
- Arenas IA, Armstrong SJ, Xu Y & Davidge ST (2005). Chronic tumor necrosis factor‐α inhibition enhances NO modulation of vascular function in estrogen‐deficient rats. Hypertension 46, 76–81. - PubMed
-
- Asai K, Kudej RK, Shen YT, Yang GP, Takagi G, Kudej AB, Geng YJ, Sato N, Nazareno JB, Vatner DE, Natividad F, Bishop SP & Vatner SF (2000). Peripheral vascular endothelial dysfunction and apoptosis in old monkeys. Arterioscler Thromb Vasc Biol 20, 1493–1499. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous