

Dynamic corticostriatal activity biases social bonding in monogamous female prairie voles
- PMID: 28562592
- PMCID: PMC5499998
- DOI: 10.1038/nature22381
Dynamic corticostriatal activity biases social bonding in monogamous female prairie voles
Abstract
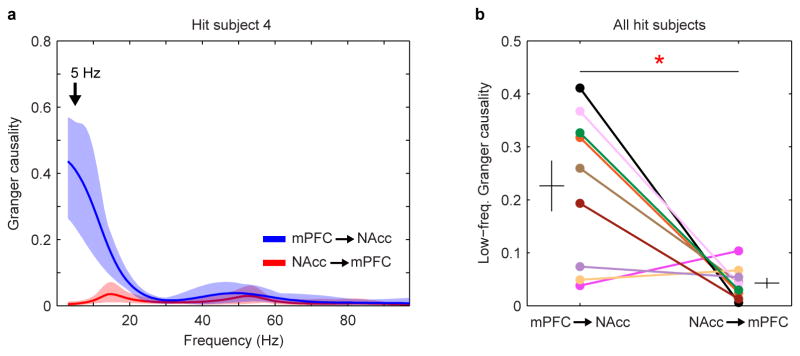
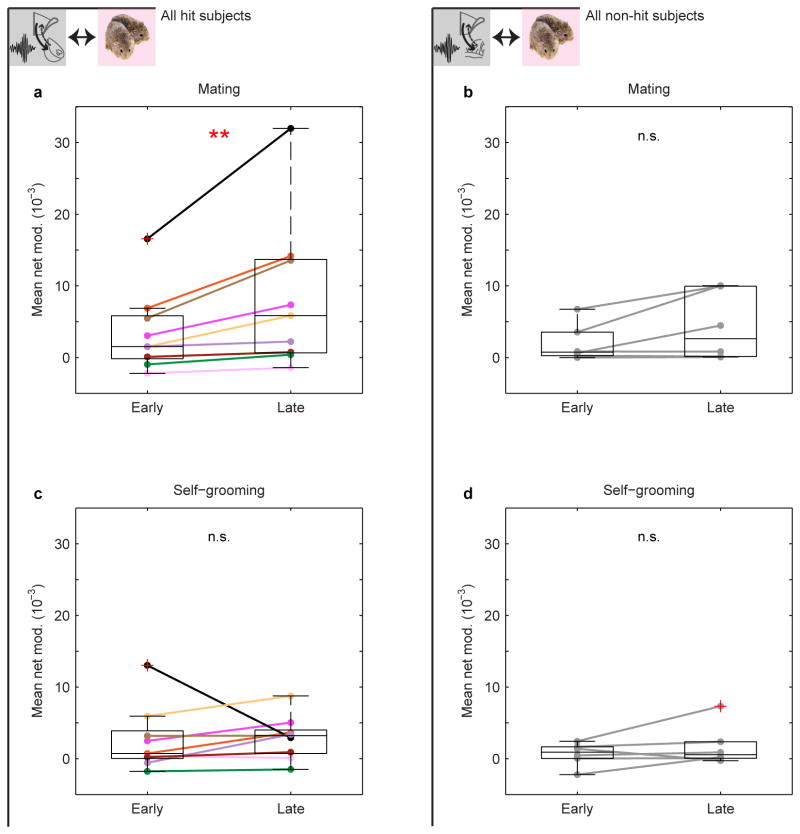
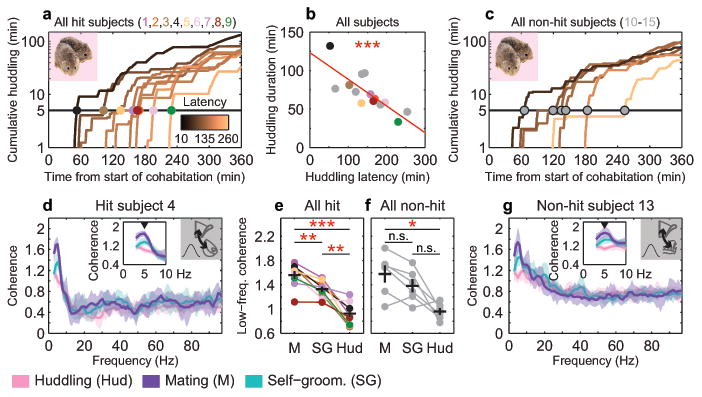
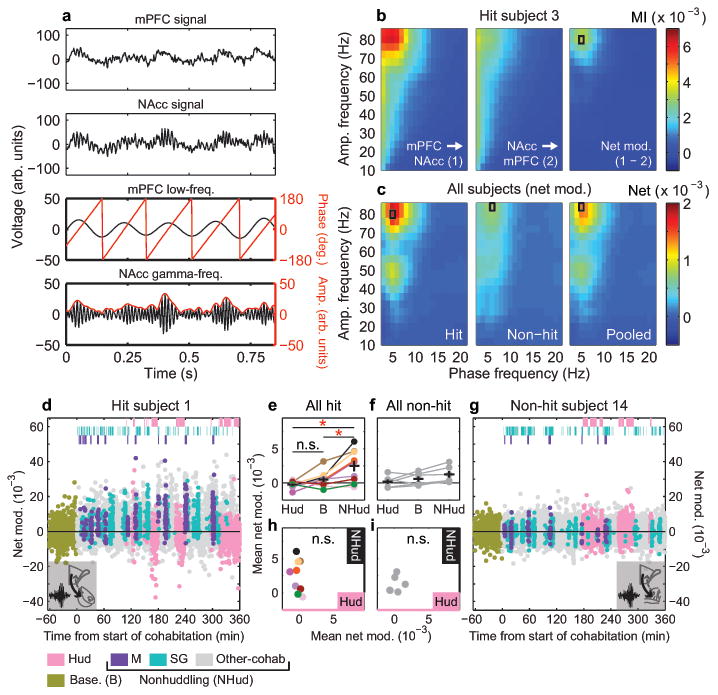
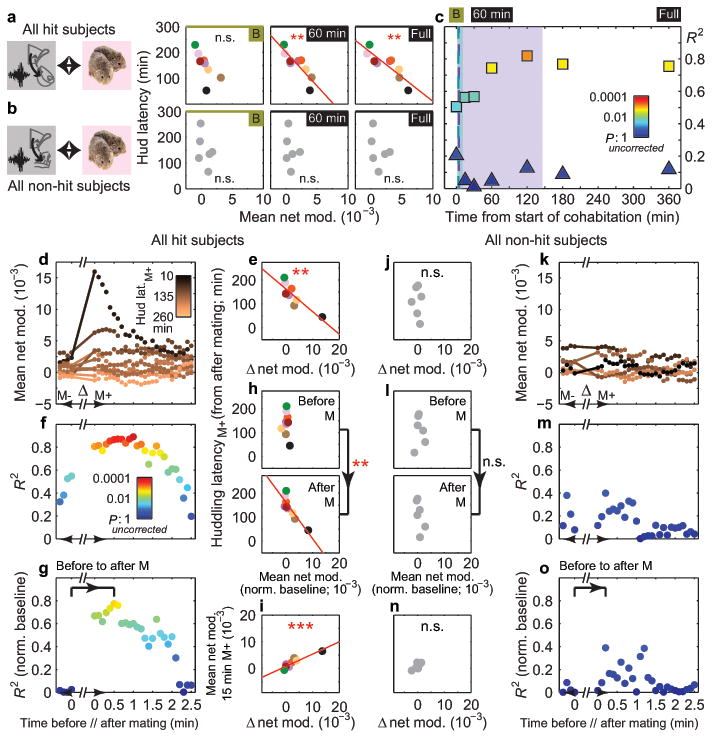
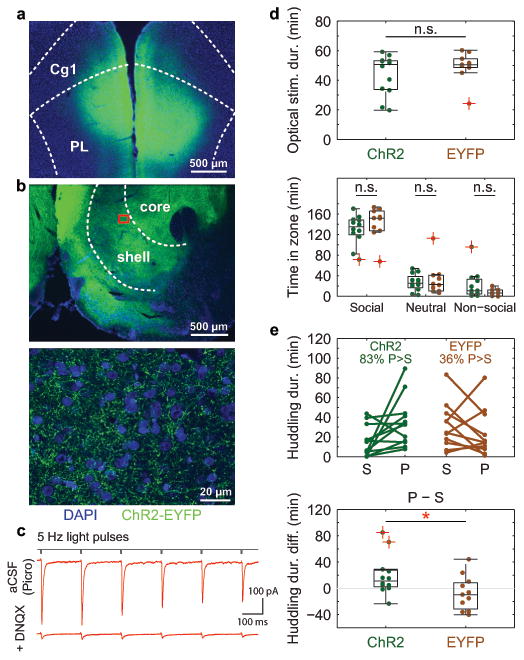
Adult pair bonding involves dramatic changes in the perception and valuation of another individual. One key change is that partners come to reliably activate the brain's reward system, although the precise neural mechanisms by which partners become rewarding during sociosexual interactions leading to a bond remain unclear. Here we show, using a prairie vole (Microtus ochrogaster) model of social bonding, how a functional circuit from the medial prefrontal cortex to nucleus accumbens is dynamically modulated to enhance females' affiliative behaviour towards a partner. Individual variation in the strength of this functional connectivity, particularly after the first mating encounter, predicts how quickly animals begin affiliative huddling with their partner. Rhythmically activating this circuit in a social context without mating biases later preference towards a partner, indicating that this circuit's activity is not just correlated with how quickly animals become affiliative but causally accelerates it. These results provide the first dynamic view of corticostriatal activity during bond formation, revealing how social interactions can recruit brain reward systems to drive changes in affiliative behaviour.
Figures
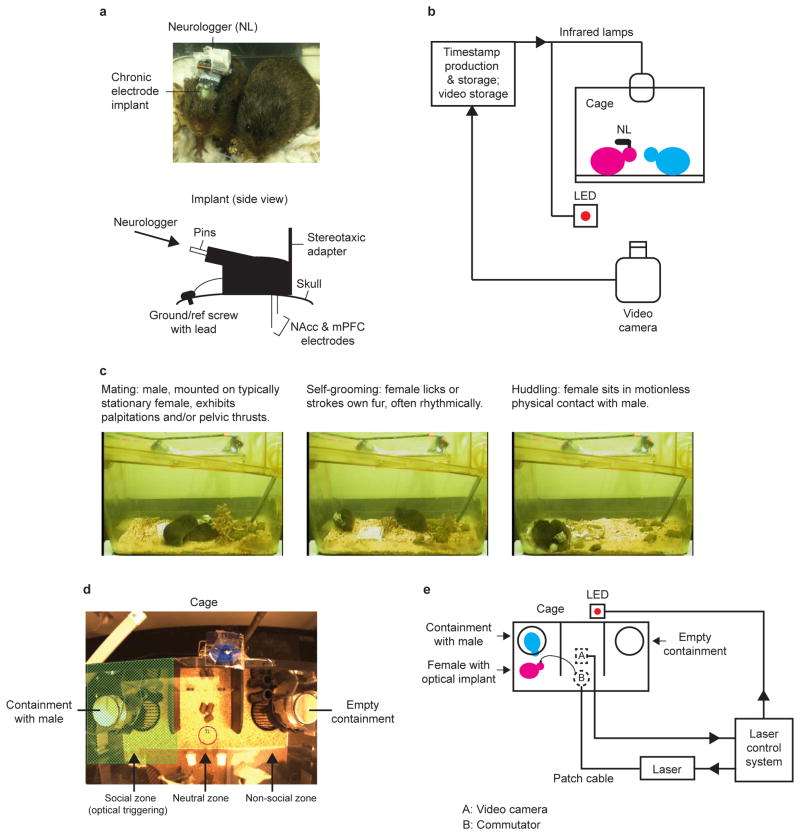
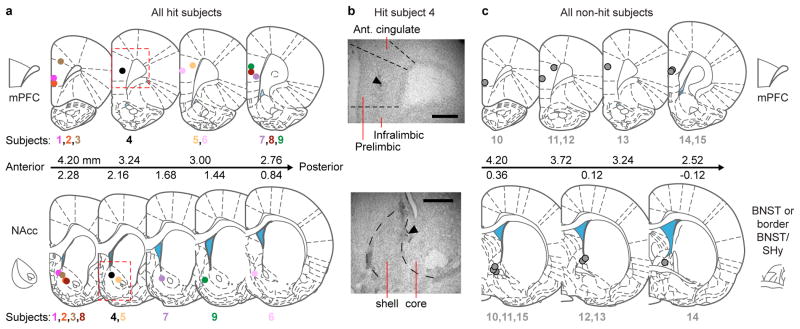
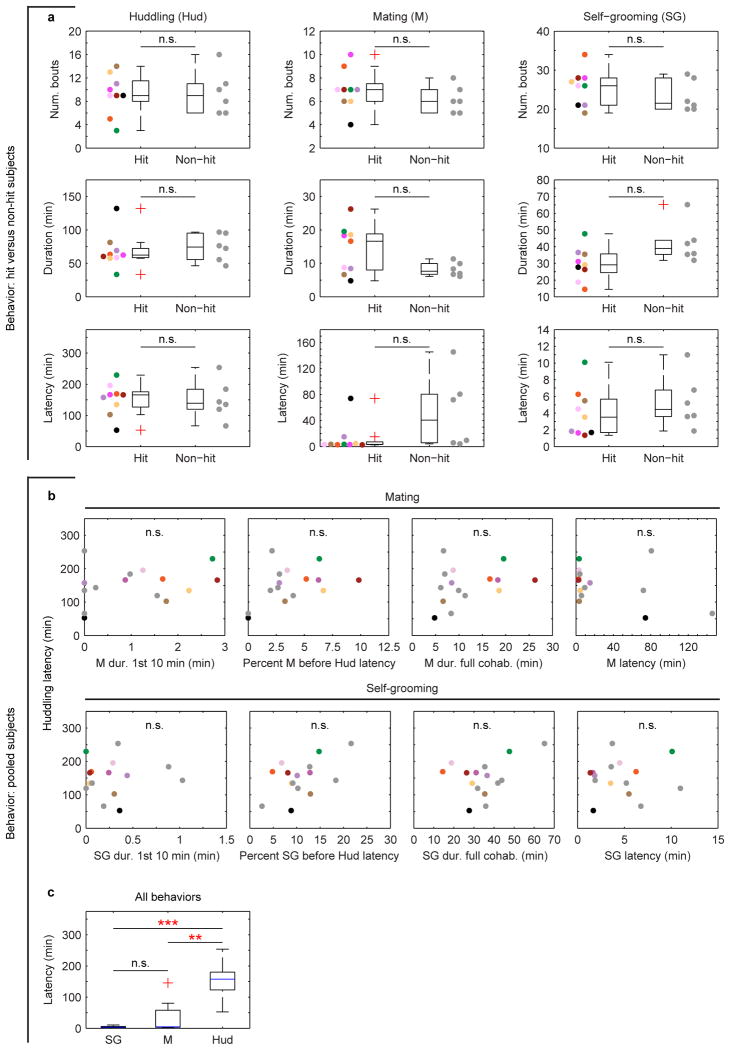
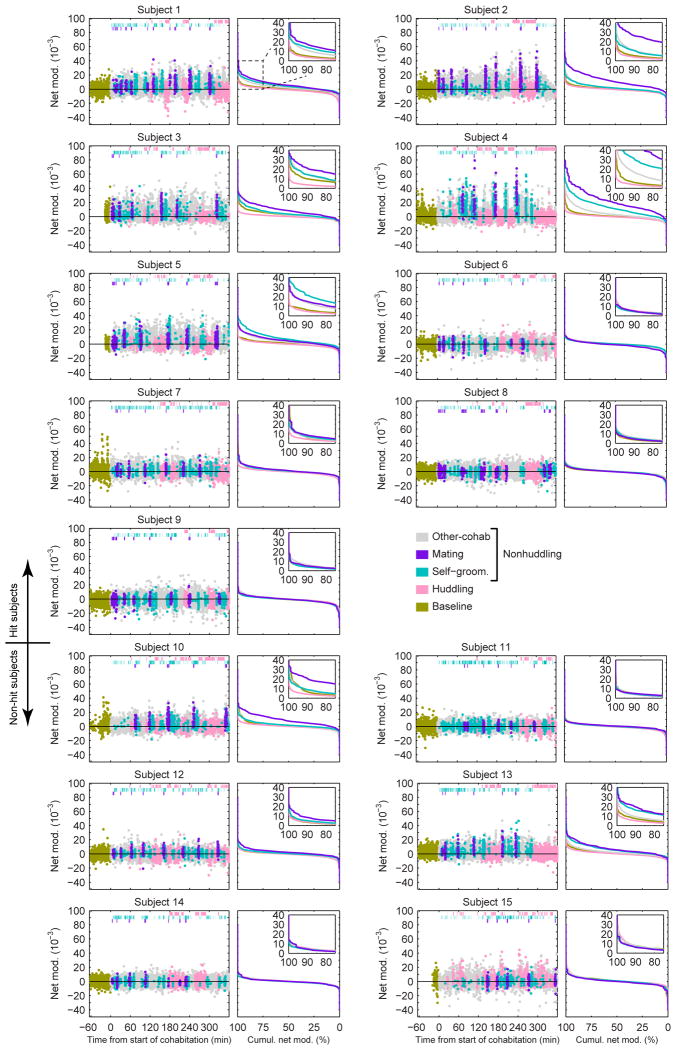
References
-
- Hazan C, Shaver P. Romantic love conceptualized as an attachment process. J Pers Soc Psychol. 1987;52:511–524. - PubMed
-
- Bartels A, Zeki S. The neural basis of romantic love. Neuroreport. 2000;11:3829–3834. - PubMed
-
- Young LJ, Lim MM, Gingrich B, Insel TR. Cellular mechanisms of social attachment. Horm Behav. 2001;40:133–138. - PubMed
-
- Aragona BJ, et al. Nucleus accumbens dopamine differentially mediates the formation and maintenance of monogamous pair bonds. Nat Neurosci. 2006;9:133–139. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
