

Microtubule-assisted altered trafficking of astrocytic gap junction protein connexin 43 is associated with depletion of connexin 47 during mouse hepatitis virus infection
- PMID: 28566289
- PMCID: PMC5592656
- DOI: 10.1074/jbc.M117.786491
Microtubule-assisted altered trafficking of astrocytic gap junction protein connexin 43 is associated with depletion of connexin 47 during mouse hepatitis virus infection
Abstract
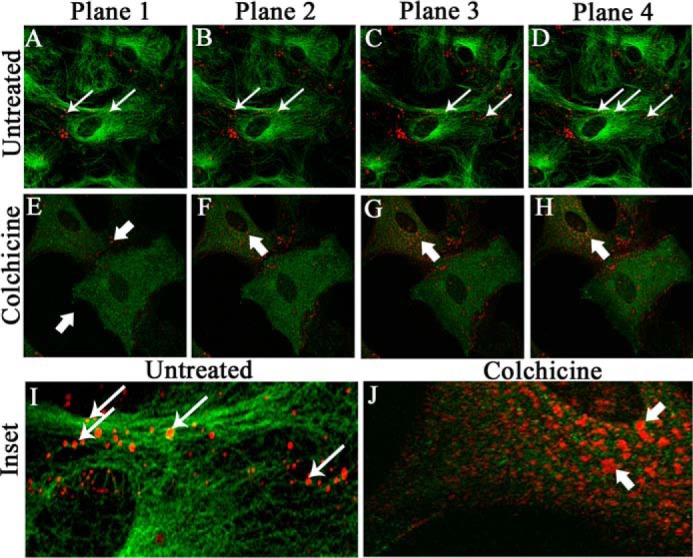
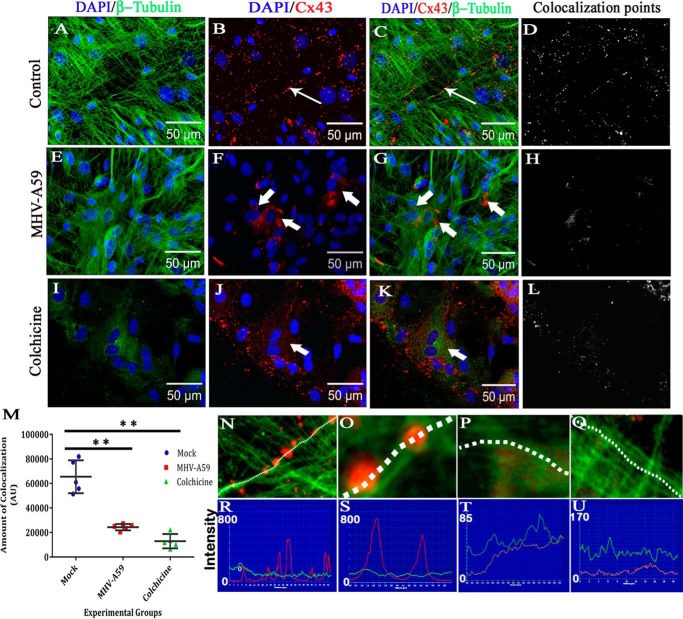
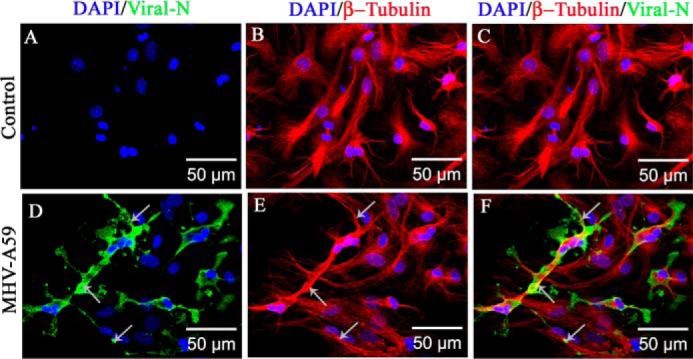
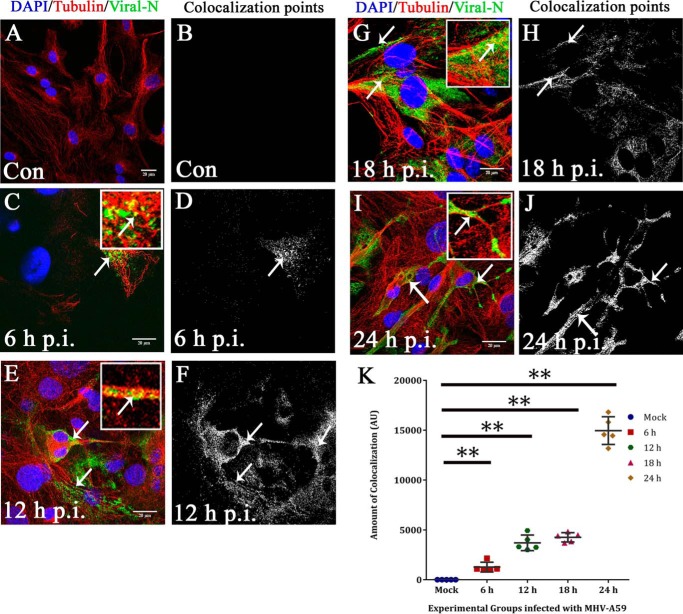
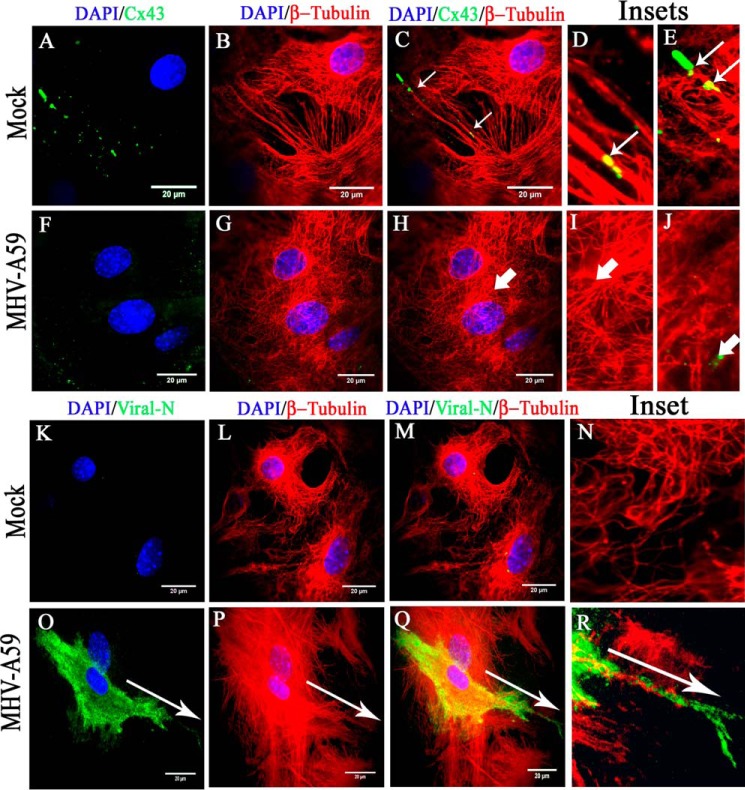
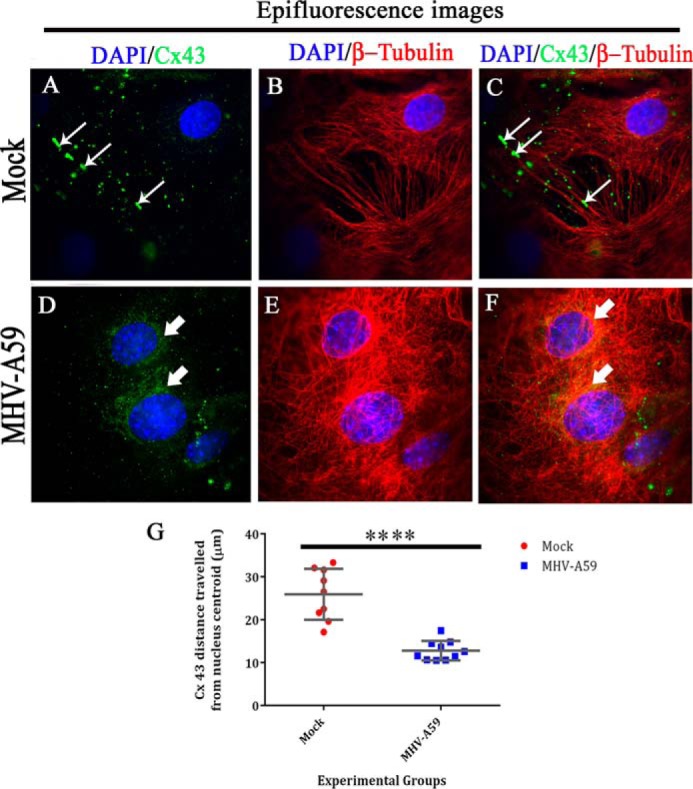
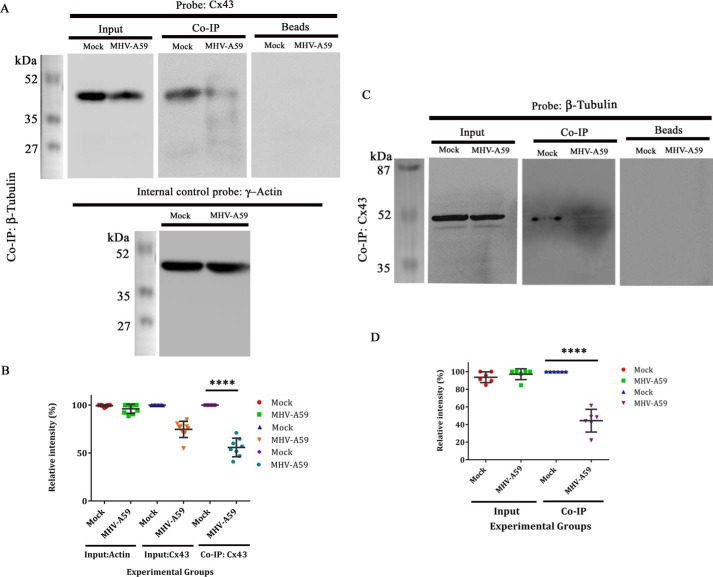
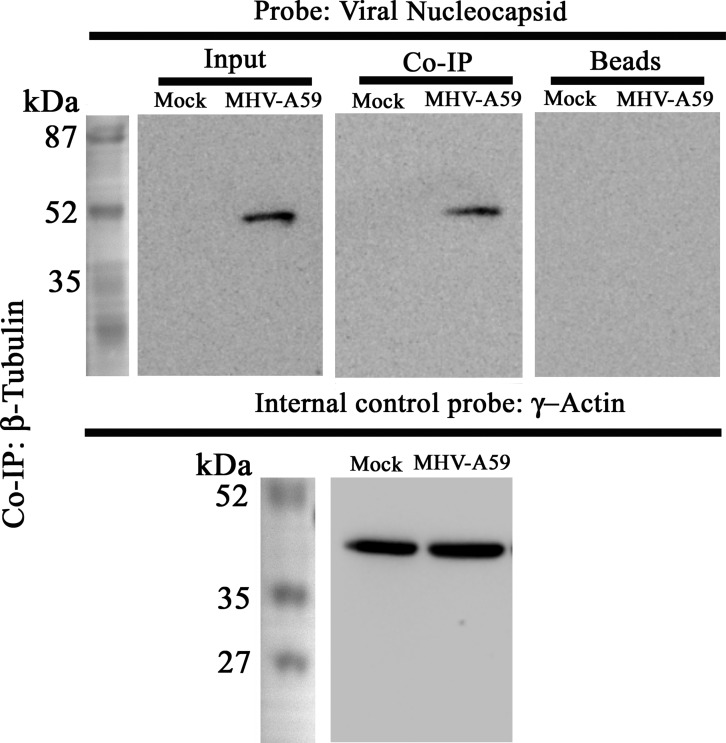
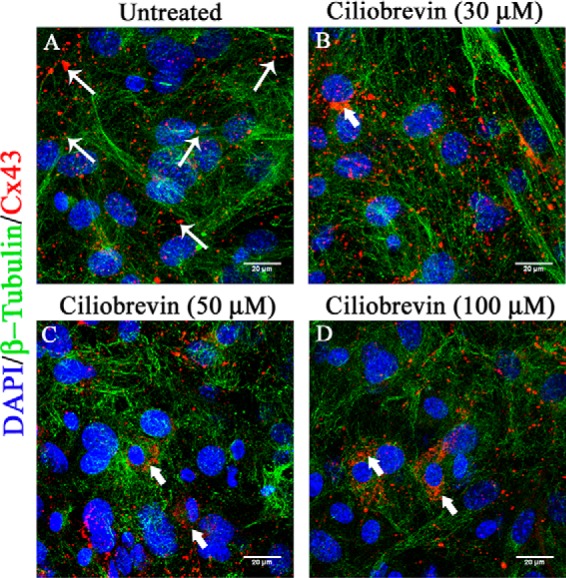
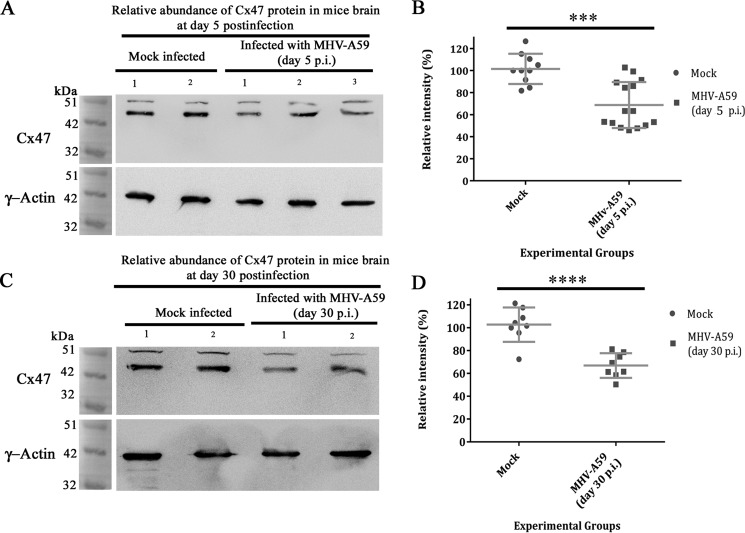
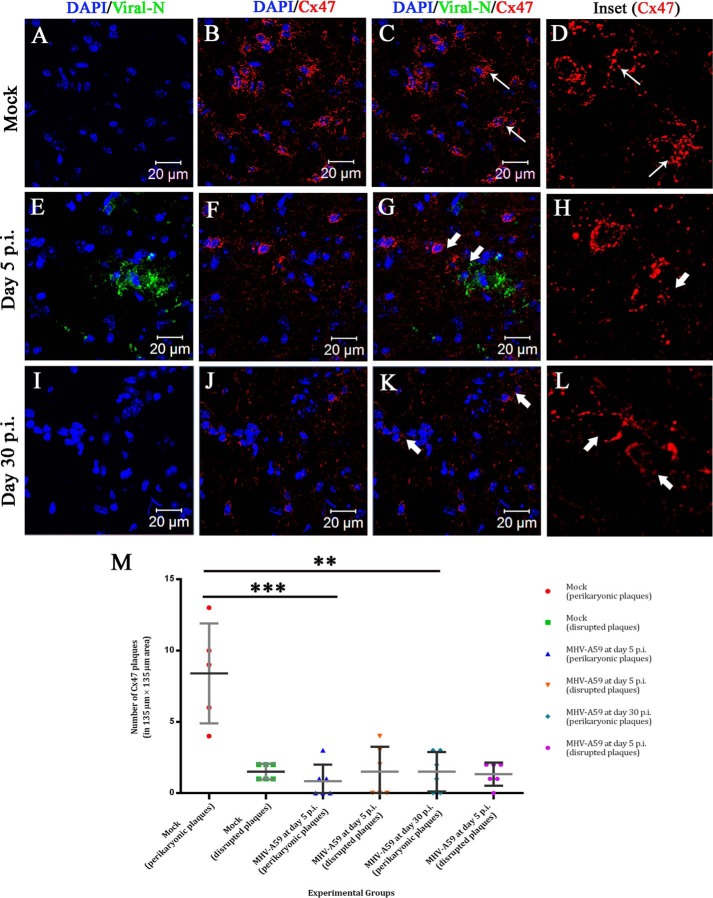
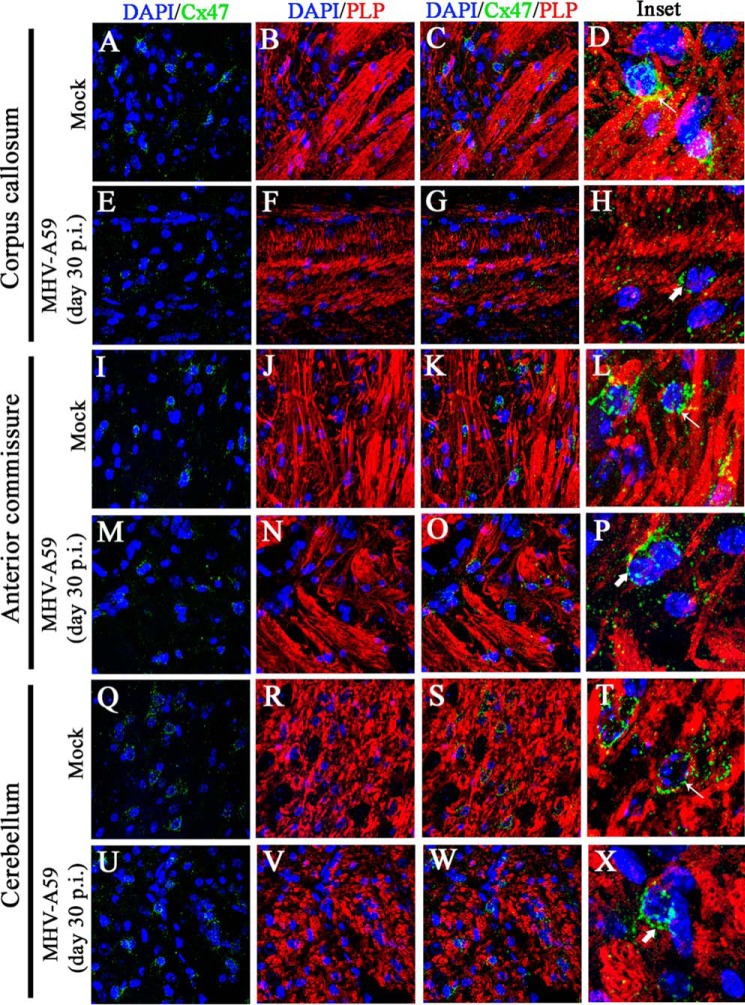
Gap junctions (GJs) are important for maintenance of CNS homeostasis. GJ proteins, connexin 43 (Cx43) and connexin 47 (Cx47), play a crucial role in production and maintenance of CNS myelin. Cx43 is mainly expressed by astrocytes in the CNS and forms gap junction intercellular communications between astrocytes-astrocytes (Cx43-Cx43) and between astrocytes-oligodendrocytes (Cx43-Cx47). Mutations of these connexin (Cx) proteins cause dysmyelinating diseases in humans. Previously, it has been shown that Cx43 localization and expression is altered due to mouse hepatitis virus (MHV)-A59 infection both in vivo and in vitro; however, its mechanism and association with loss of myelin protein was not elaborated. Thus, we explored potential mechanisms by which MHV-A59 infection alters Cx43 localization and examined the effects of viral infection on Cx47 expression and its association with loss of the myelin marker proteolipid protein. Immunofluorescence and total internal reflection fluorescence microscopy confirmed that MHV-A59 used microtubules (MTs) as a conduit to reach the cell surface and restricted MT-mediated Cx43 delivery to the cell membrane. Co-immunoprecipitation experiments demonstrated that Cx43-β-tubulin molecular interaction was depleted due to protein-protein interaction between viral particles and MTs. During acute MHV-A59 infection, oligodendrocytic Cx47, which is mainly stabilized by Cx43 in vivo, was down-regulated, and its characteristic staining remained disrupted even at chronic phase. The loss of Cx47 was associated with loss of proteolipid protein at the chronic stage of MHV-A59 infection.
Keywords: astrocyte; connexin; gap junction; hepatitis virus; microtubule; myelin; oligodendrocyte; tubulin.
© 2017 by The American Society for Biochemistry and Molecular Biology, Inc.
Conflict of interest statement
The authors declare that they have no conflicts of interest with the contents of this article
Figures
References
-
- Das Sarma J., Wang F., and Koval M. (2002) Targeted gap junction protein constructs reveal connexin-specific differences in oligomerization. J. Biol. Chem. 277, 20911–20918 - PubMed
-
- Charles A. C., Merrill J. E., Dirksen E. R., and Sanderson M. J. (1991) Intercellular signaling in glial cells: calcium waves and oscillations in response to mechanical stimulation and glutamate. Neuron 6, 983–992 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
