

Structural Insight into Nucleoprotein Conformation Change Chaperoned by VP35 Peptide in Marburg Virus
- PMID: 28566377
- PMCID: PMC5533893
- DOI: 10.1128/JVI.00825-17
Structural Insight into Nucleoprotein Conformation Change Chaperoned by VP35 Peptide in Marburg Virus
Abstract
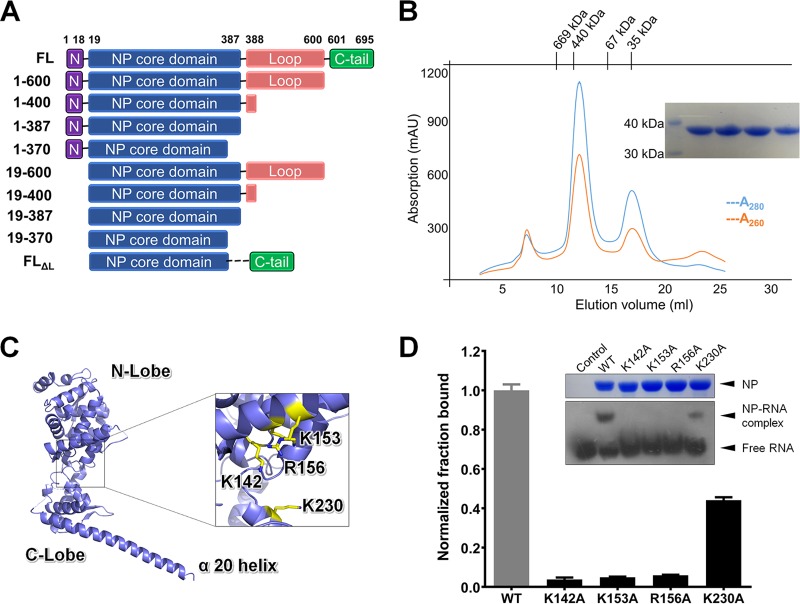
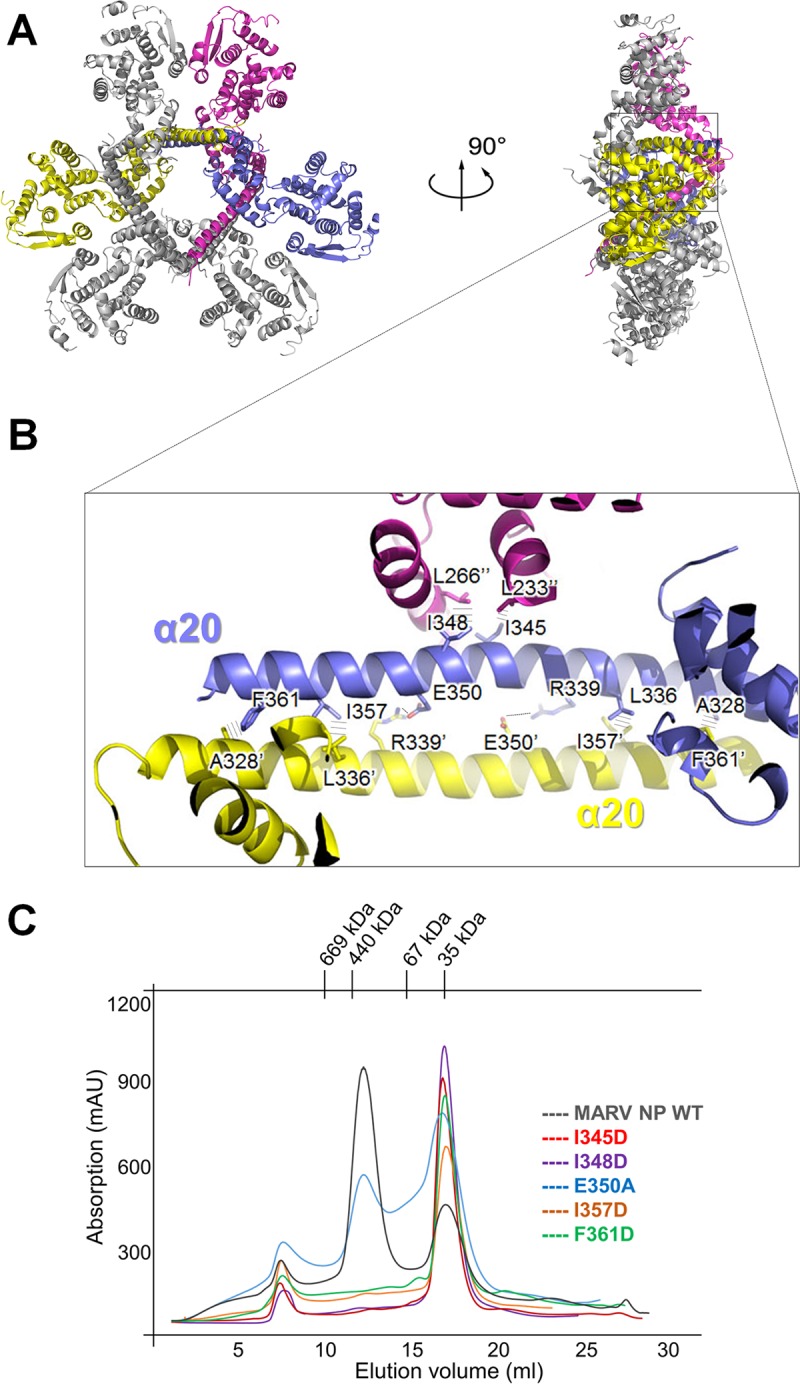
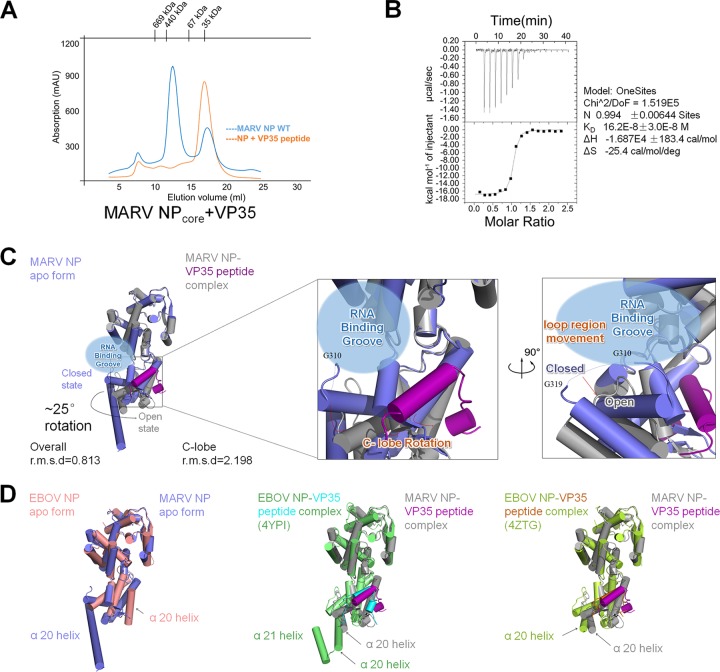
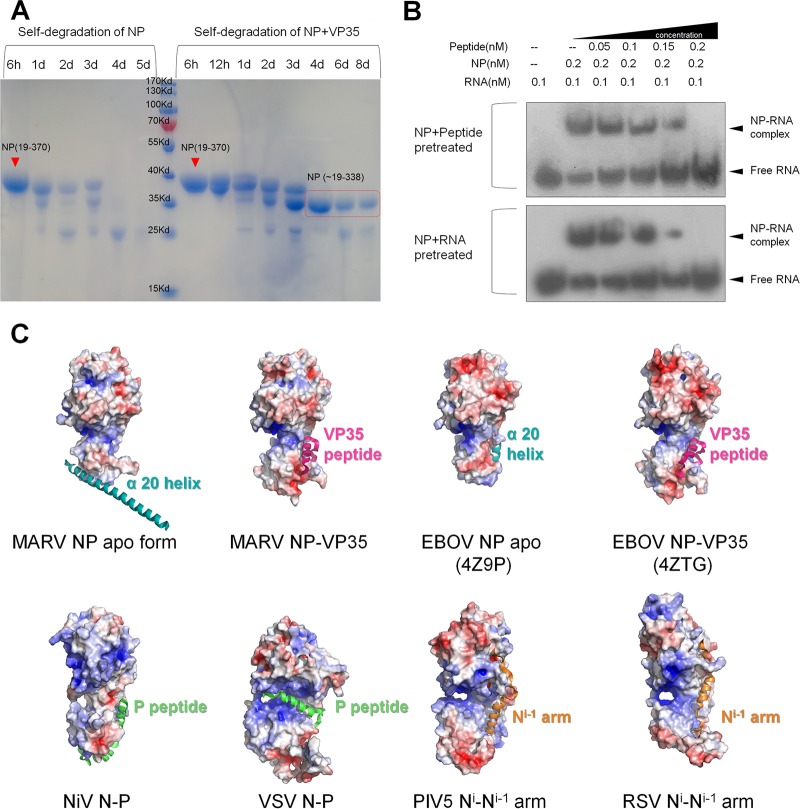
Marburg virus (MARV) encodes a nucleoprotein (NP) to encapsidate its genome by oligomerization and form a ribonucleoprotein complex (RNP). According to previous investigation on nonsegmented negative-sense RNA viruses (nsNSV), the newly synthesized NPs must be prevented from indiscriminately binding to noncognate RNAs. During the viral RNA synthesis process, the RNPs undergo a transition from an RNA-bound form to a template-free form, to open access for the interaction between the viral polymerase and the RNA template. In filoviruses, this transition is regulated by VP35 peptide and other viral components. To further understand the dynamic process of filovirus RNP formation, we report here the structure of MARV NPcore, both in the apo form and in the VP35 peptide-chaperoned form. These structures reveal a typical bilobed structure, with a positive-charged RNA binding groove between two lobes. In the apo form, the MARV NP exists in an interesting hexameric state formed by the hydrophobic interaction within the long helix of the NPcore C-terminal region, which shows high structural flexibility among filoviruses and may imply critical function during RNP formation. Moreover, the VP35 peptide-chaperoned NPcore remains in a monomeric state and completely loses its affinity for single-stranded RNA (ssRNA). The structural comparison reveals that the RNA binding groove undergoes a transition from closed state to open state, chaperoned by VP35 peptide, thus preventing the interaction for viral RNA. Our investigation provides considerable structural insight into the filovirus RNP working mechanism and may support the development of antiviral therapies targeting the RNP formation of filovirus.IMPORTANCE Marburg virus is one of the most dangerous viruses, with high morbidity and mortality. A recent outbreak in Angola in 2005 caused the deaths of 272 persons. NP is one of the most essential proteins, as it encapsidates and protects the whole virus genome simultaneously with self-assembly oligomerization. Here we report the structures of MARV NPcore in two different forms. In the MARV NP apo form, we identify an interesting hexamer formed by hydrophobic interaction within a long helix, which is highly conserved and flexible among filoviruses and may indicate its critical function during the virus RNP formation. Moreover, the structural comparison with the NP-VP35 peptide complex reveals a structural transition chaperoned by VP35, in which the RNA binding groove undergoes a transition from closed state to open state. Finally, we discussed the high conservation and critical role of the VP35 binding pocket and its potential use for therapeutic development.
Keywords: Marburg virus; assembly mechanism; crystal structure; filovirus; nucleoprotein.
Copyright © 2017 American Society for Microbiology.
Figures
Similar articles
-
Crystal Structure of the Marburg Virus Nucleoprotein Core Domain Chaperoned by a VP35 Peptide Reveals a Conserved Drug Target for Filovirus.J Virol. 2017 Aug 24;91(18):e00996-17. doi: 10.1128/JVI.00996-17. Print 2017 Sep 15. J Virol. 2017. PMID: 28659479 Free PMC article.
-
Sudan Ebolavirus VP35-NP Crystal Structure Reveals a Potential Target for Pan-Filovirus Treatment.mBio. 2019 Jul 23;10(4):e00734-19. doi: 10.1128/mBio.00734-19. mBio. 2019. PMID: 31337716 Free PMC article.
-
Crystal Structure of the Marburg Virus VP35 Oligomerization Domain.J Virol. 2017 Jan 3;91(2):e01085-16. doi: 10.1128/JVI.01085-16. Print 2017 Jan 15. J Virol. 2017. PMID: 27847355 Free PMC article.
-
Filovirus helical nucleocapsid structures.Microscopy (Oxf). 2023 Jun 8;72(3):178-190. doi: 10.1093/jmicro/dfac049. Microscopy (Oxf). 2023. PMID: 36242583 Review.
-
Evasion of interferon responses by Ebola and Marburg viruses.J Interferon Cytokine Res. 2009 Sep;29(9):511-20. doi: 10.1089/jir.2009.0076. J Interferon Cytokine Res. 2009. PMID: 19694547 Free PMC article. Review.
Cited by
-
Cryoelectron microscopic structure of the nucleoprotein-RNA complex of the European filovirus, Lloviu virus.PNAS Nexus. 2023 Apr 6;2(4):pgad120. doi: 10.1093/pnasnexus/pgad120. eCollection 2023 Apr. PNAS Nexus. 2023. PMID: 37124400 Free PMC article.
-
Structure and Dynamics of the Unassembled Nucleoprotein of Rabies Virus in Complex with Its Phosphoprotein Chaperone Module.Viruses. 2022 Dec 16;14(12):2813. doi: 10.3390/v14122813. Viruses. 2022. PMID: 36560817 Free PMC article.
-
Zinc and Copper Ions Differentially Regulate Prion-Like Phase Separation Dynamics of Pan-Virus Nucleocapsid Biomolecular Condensates.Viruses. 2020 Oct 18;12(10):1179. doi: 10.3390/v12101179. Viruses. 2020. PMID: 33081049 Free PMC article.
-
Emergence of Marburg virus: a global perspective on fatal outbreaks and clinical challenges.Front Microbiol. 2023 Sep 13;14:1239079. doi: 10.3389/fmicb.2023.1239079. eCollection 2023. Front Microbiol. 2023. PMID: 37771708 Free PMC article. Review.
-
How does the polymerase of non-segmented negative strand RNA viruses commit to transcription or genome replication?J Virol. 2024 Aug 20;98(8):e0033224. doi: 10.1128/jvi.00332-24. Epub 2024 Jul 30. J Virol. 2024. PMID: 39078194 Free PMC article. Review.
References
-
- Bausch DG, Nichol ST, Muyembe-Tamfum JJ, Borchert M, Rollin PE, Sleurs H, Campbell P, Tshioko FK, Roth C, Colebunders R, Pirard P, Mardel S, Olinda LA, Zeller H, Tshomba A, Kulidri A, Libande ML, Mulangu S, Formenty P, Grein T, Leirs H, Braack L, Ksiazek T, Zaki S, Bowen MD, Smit SB, Leman PA, Burt FJ, Kemp A, Swanepoel R, International Scientific and Technical Committee for Marburg Hemorrhagic Fever Control in the Democratic Republic of the Congo. 2006. Marburg hemorrhagic fever associated with multiple genetic lineages of virus. N Engl J Med 355:909–919. doi:10.1056/NEJMoa051465. - DOI - PubMed
-
- Towner JS, Khristova ML, Sealy TK, Vincent MJ, Erickson BR, Bawiec DA, Hartman AL, Comer JA, Zaki SR, Stroher U, Gomes da Silva F, del Castillo F, Rollin PE, Ksiazek TG, Nichol ST. 2006. Marburgvirus genomics and association with a large hemorrhagic fever outbreak in Angola. J Virol 80:6497–6516. doi:10.1128/JVI.00069-06. - DOI - PMC - PubMed
-
- Kuhn JH, Becker S, Ebihara H, Geisbert TW, Johnson KM, Kawaoka Y, Lipkin WI, Negredo AI, Netesov SV, Nichol ST, Palacios G, Peters CJ, Tenorio A, Volchkov VE, Jahrling PB. 2010. Proposal for a revised taxonomy of the family Filoviridae: classification, names of taxa and viruses, and virus abbreviations. Arch Virol 155:2083–2103. doi:10.1007/s00705-010-0814-x. - DOI - PMC - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
