

Building early-larval sexing systems for genetic control of the Australian sheep blow fly Lucilia cuprina using two constitutive promoters
- PMID: 28566730
- PMCID: PMC5451413
- DOI: 10.1038/s41598-017-02763-4
Building early-larval sexing systems for genetic control of the Australian sheep blow fly Lucilia cuprina using two constitutive promoters
Abstract
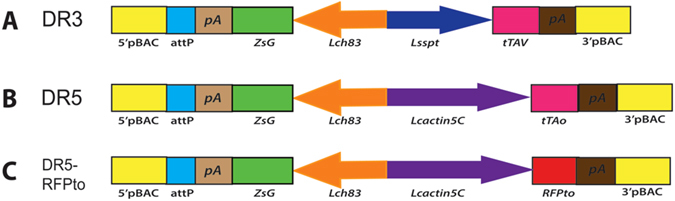
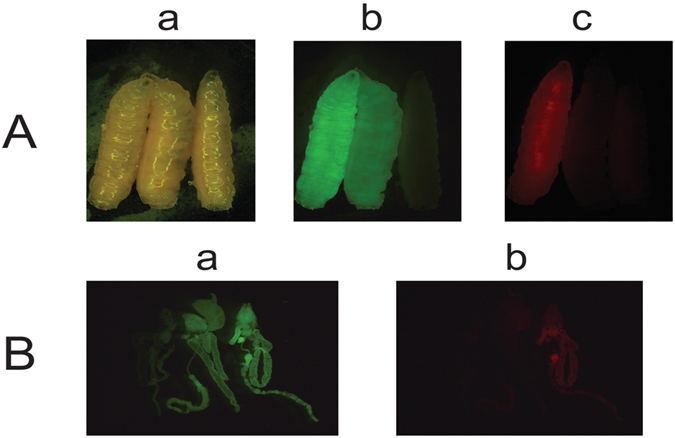
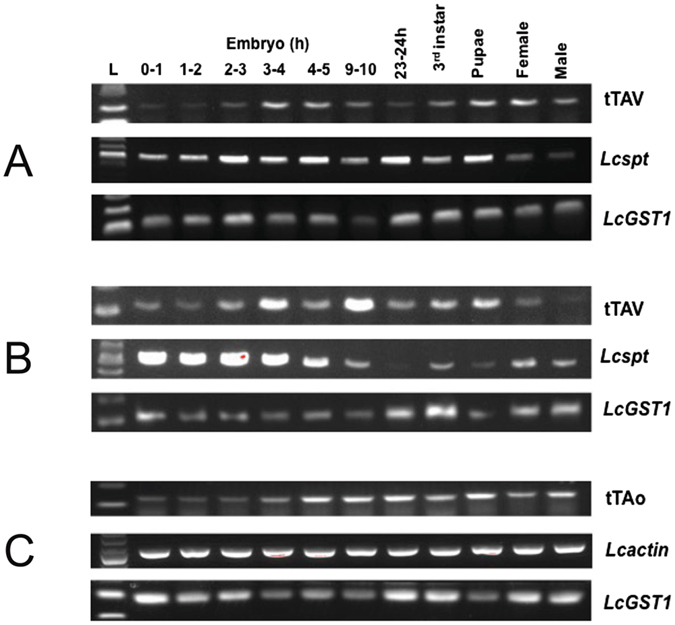
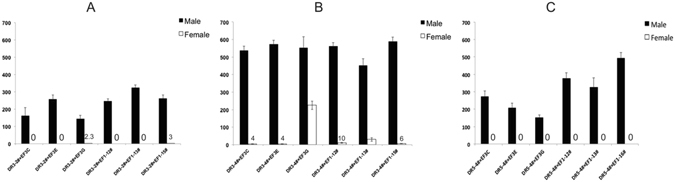
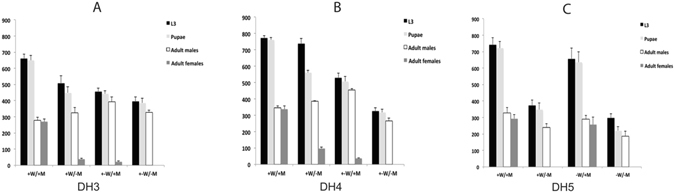
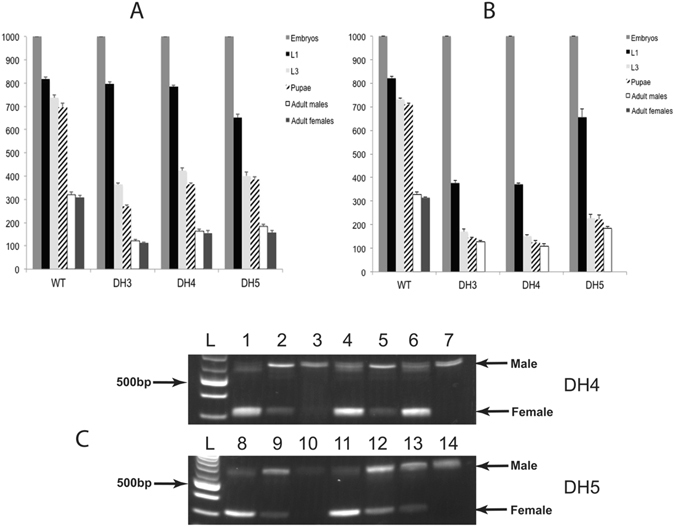
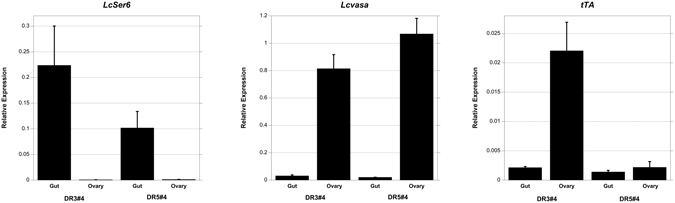
Transgenic sexing strains (TSS) that carry conditional female lethal genes are advantageous for genetic control programs based on the sterile insect technique (SIT). It is desirable if females die early in development as larval diet is a major cost for mass production facilities. This can be achieved by using a gene promoter that is only active in embryos to drive expression of the tetracycline transactivator (tTA), the transcription factor commonly used in two-component TSS. While an embryo-specific promoter is ideal it may not be essential for assembling an effective TSS as tTA can be repressed by addition of tetracycline to the diet at larval and/or adult stages. Here we have investigated this idea by isolating and employing the promoters from the Lucilia spitting image and actin 5C genes to drive tTA expression in embryos and later stages. L. cuprina TSS with the tTA drivers and tTA-regulated tetO-Lshid effectors produced only females when raised on a limited tetracycline diet. The Lshid transgene contains a sex-specific intron and as a consequence only females produce LsHID protein. TSS females died at early larval stages, which makes the lines advantageous for an SIT program.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures
Similar articles
-
A transgenic embryonic sexing system for the Australian sheep blow fly Lucilia cuprina.Sci Rep. 2015 Nov 5;5:16090. doi: 10.1038/srep16090. Sci Rep. 2015. PMID: 26537204 Free PMC article.
-
Building a transgenic sexing strain for genetic control of the Australian sheep blow fly Lucilia cuprina using two lethal effectors.BMC Genet. 2020 Dec 18;21(Suppl 2):141. doi: 10.1186/s12863-020-00947-y. BMC Genet. 2020. PMID: 33339506 Free PMC article.
-
Improved transgenic sexing strains for genetic control of the Australian sheep blow fly Lucilia cuprina using embryo-specific gene promoters.Mol Genet Genomics. 2020 Mar;295(2):287-298. doi: 10.1007/s00438-019-01622-3. Epub 2019 Nov 12. Mol Genet Genomics. 2020. PMID: 31720776
-
Development and evaluation of male-only strains of the Australian sheep blowfly, Lucilia cuprina.BMC Genet. 2014;15 Suppl 2(Suppl 2):S3. doi: 10.1186/1471-2156-15-S2-S3. Epub 2014 Dec 1. BMC Genet. 2014. PMID: 25472415 Free PMC article. Review.
-
Enhancing the stability and ecological safety of mass-reared transgenic strains for field release by redundant conditional lethality systems.Insect Sci. 2016 Apr;23(2):225-34. doi: 10.1111/1744-7917.12245. Epub 2015 Oct 26. Insect Sci. 2016. PMID: 26097098 Review.
Cited by
-
Tetracycline has no long-term effects on gut physiology and microbiome of the new world screwworm, Cochliomyia hominivorax, which has positive implications for transgenic male-only rearing systems.G3 (Bethesda). 2025 May 8;15(5):jkaf058. doi: 10.1093/g3journal/jkaf058. G3 (Bethesda). 2025. PMID: 40130547 Free PMC article.
-
Genetic and behavioral analyses suggest that larval and adult stages of Lucilia cuprina employ different sensory systems to detect rotten beef.Parasit Vectors. 2025 Jul 9;18(1):270. doi: 10.1186/s13071-025-06804-0. Parasit Vectors. 2025. PMID: 40635088 Free PMC article.
-
Effects of antibiotics on the in vitro expression of tetracycline-off constructs and the performance of Drosophila suzukii female-killing strains.Front Bioeng Biotechnol. 2023 Feb 14;11:876492. doi: 10.3389/fbioe.2023.876492. eCollection 2023. Front Bioeng Biotechnol. 2023. PMID: 36865029 Free PMC article.
-
An early female lethal system of the New World screwworm, Cochliomyia hominivorax, for biotechnology-enhanced SIT.BMC Genet. 2020 Dec 18;21(Suppl 2):143. doi: 10.1186/s12863-020-00948-x. BMC Genet. 2020. PMID: 33339501 Free PMC article.
-
Genetic breakdown of a Tet-off conditional lethality system for insect population control.Nat Commun. 2020 Jun 18;11(1):3095. doi: 10.1038/s41467-020-16807-3. Nat Commun. 2020. PMID: 32555259 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
