

Phospho-specific antibodies targeting the amino terminus of the human dopamine transporter
- PMID: 28571709
- PMCID: PMC5565701
- DOI: 10.1016/j.jchemneu.2017.05.003
Phospho-specific antibodies targeting the amino terminus of the human dopamine transporter
Abstract
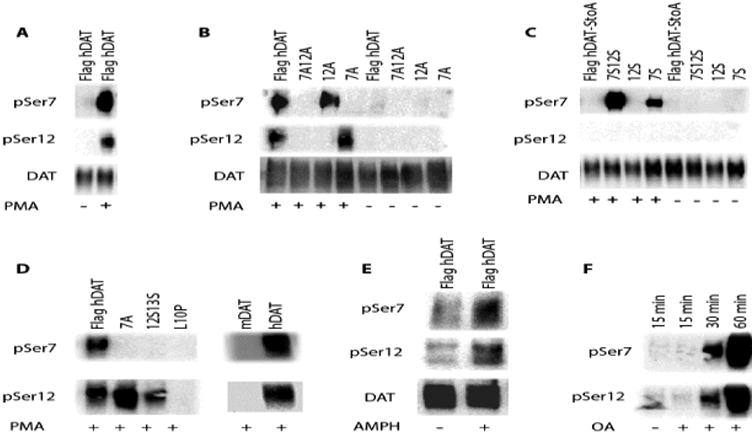
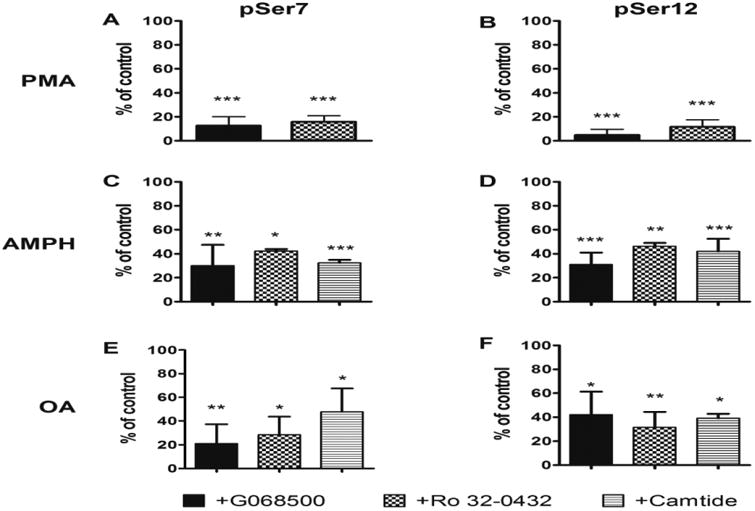
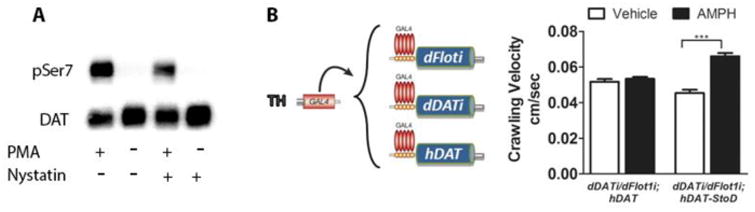
The dopamine transporter (DAT), which mediates the inactivation of released dopamine through its reuptake, is the primary molecular target for the actions of psychostimulants. An increasing number of studies support an essential role for phosphorylation of serines (Ser) in the distal amino (N) terminus of DAT in regulating its function. Still, the molecular details of the regulation of phosphorylation and its impact on function are not fully understood. To address this, we have developed and characterized two distinct phospho-antibodies that recognize human DAT when it is phosphorylated at Ser7 or Ser12. Our data show that treatment of cells with phorbol 12-myristate 13-acetate (PMA), amphetamine (AMPH) or okadaic acid (OA) leads to an increase in the phosphorylation of DAT at both residues and that these responses are dependent on the activity of protein kinase C. We also show that AMPH-induced and OA-induced phosphorylation of DAT are dependent on Ca2+/calmodulin-dependent protein kinase α. Our data further suggest that the lipid raft localization of DAT is necessary for efficient N-terminal phosphorylation and for the associated behavioral effects of AMPH, demonstrating the potential of these novel antibodies as powerful tools to study DAT regulation and function in vivo.
Keywords: Amphetamine; CamKII; Dopamine efflux; Lipid rafts; Locomotor behavior; Phosphorylation.
Copyright © 2017 Elsevier B.V. All rights reserved.
Figures
References
-
- Adkins EM, Samuvel DJ, Fog JU, Eriksen J, Jayanthi LD, Vaegter CB, Ramamoorthy S, Gether U. Membrane mobility and microdomain association of the dopamine transporter studied with fluorescence correlation spectroscopy and fluorescence recovery after photobleaching. Biochemistry. 2007;46:10484–10497. - PubMed
-
- Amara SG, Kuhar MJ. Neurotransmitter transporters: recent progress. Annu Rev Neurosci. 1993;16:73–93. - PubMed
-
- Bowton E, Saunders C, Erreger K, Sakrikar D, Matthies HJ, Sen N, Jessen T, Colbran RJ, Caron MG, Javitch JA, et al. Dysregulation of dopamine transporters via dopamine D2 autoreceptors triggers anomalous dopamine efflux associated with attention-deficit hyperactivity disorder. J Neurosci. 2010;30:6048–6057. - PMC - PubMed
-
- Bruss M, Wieland A, Bonisch H. Molecular cloning and functional expression of the mouse dopamine transporter. J Neural Transm (Vienna) 1999;106:657–662. - PubMed
-
- Cervinski MA, Foster JD, Vaughan RA. Psychoactive substrates stimulate dopamine transporter phosphorylation and down-regulation by cocaine-sensitive and protein kinase C-dependent mechanisms. The Journal of biological chemistry. 2005;280:40442–40449. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
