

LTA4H regulates cell cycle and skin carcinogenesis
- PMID: 28575166
- PMCID: PMC6248358
- DOI: 10.1093/carcin/bgx049
LTA4H regulates cell cycle and skin carcinogenesis
Abstract
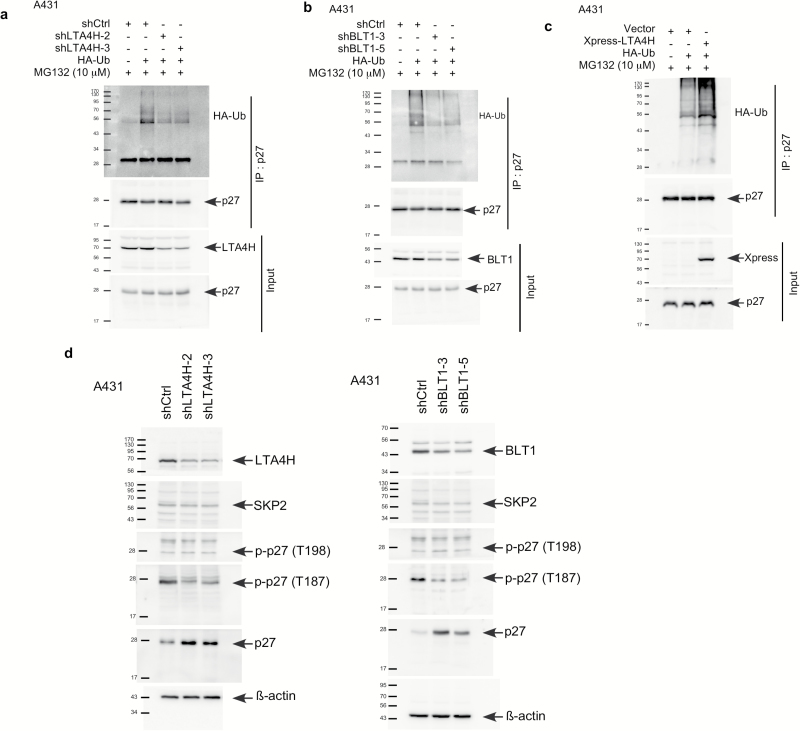
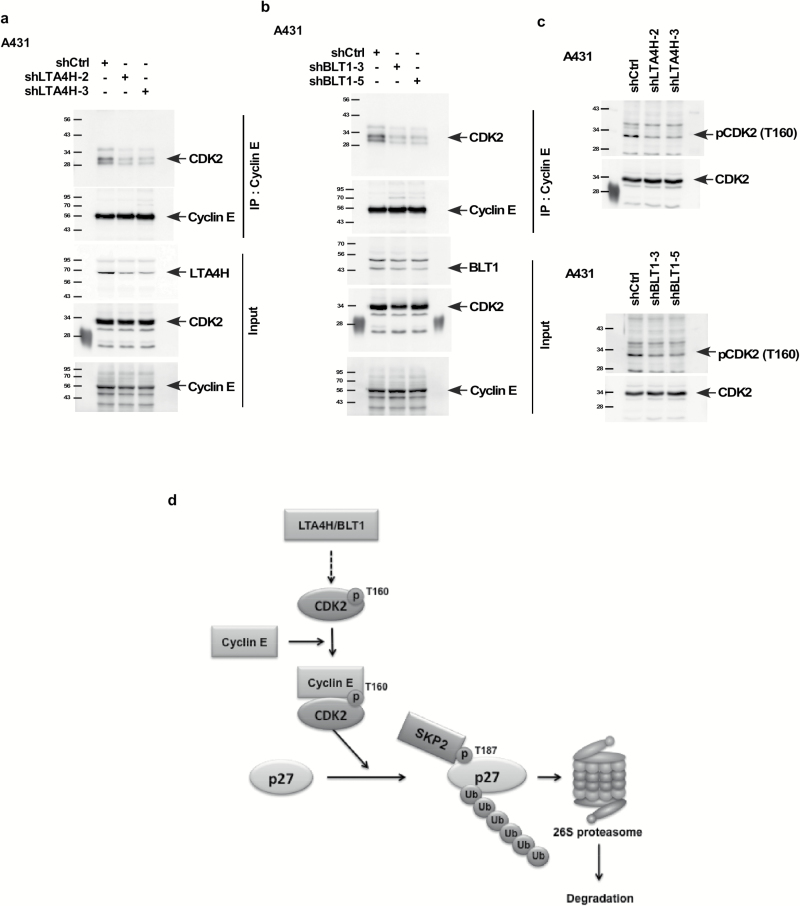
Leukotriene A4 hydrolase (LTA4H), a bifunctional zinc metallo-enzyme, is reportedly overexpressed in several human cancers. Our group has focused on LTA4H as a potential target for cancer prevention and/or therapy. In the present study, we report that LTA4H is a key regulator of cell cycle at the G0/G1 phase acting by negatively regulating p27 expression in skin cancer. We found that LTA4H is overexpressed in human skin cancer tissue. Knocking out LTA4H significantly reduced skin cancer development in the 7,12-dimethylbenz(a)anthracene (DMBA)-initiated/12-O-tetradecanoylphorbol-13-acetate (TPA)-promoted two-stage skin cancer mouse model. LTA4H depletion dramatically decreased anchorage-dependent and -independent skin cancer cell growth by inducing cell cycle arrest at the G0/G1 phase. Moreover, our findings showed that depletion of LTA4H enhanced p27 protein stability, which was associated with decreased phosphorylation of CDK2 at Thr160 and inhibition of the CDK2/cyclin E complex, resulting in down-regulated p27 ubiquitination. These findings indicate that LTA4H is critical for skin carcinogenesis and is an important mediator of cell cycle and the data begin to clarify the mechanisms of LTA4H's role in cancer development.
© The Author 2017. Published by Oxford University Press. All rights reserved. For Permissions, please email: journals.permissions@oup.com.
Figures
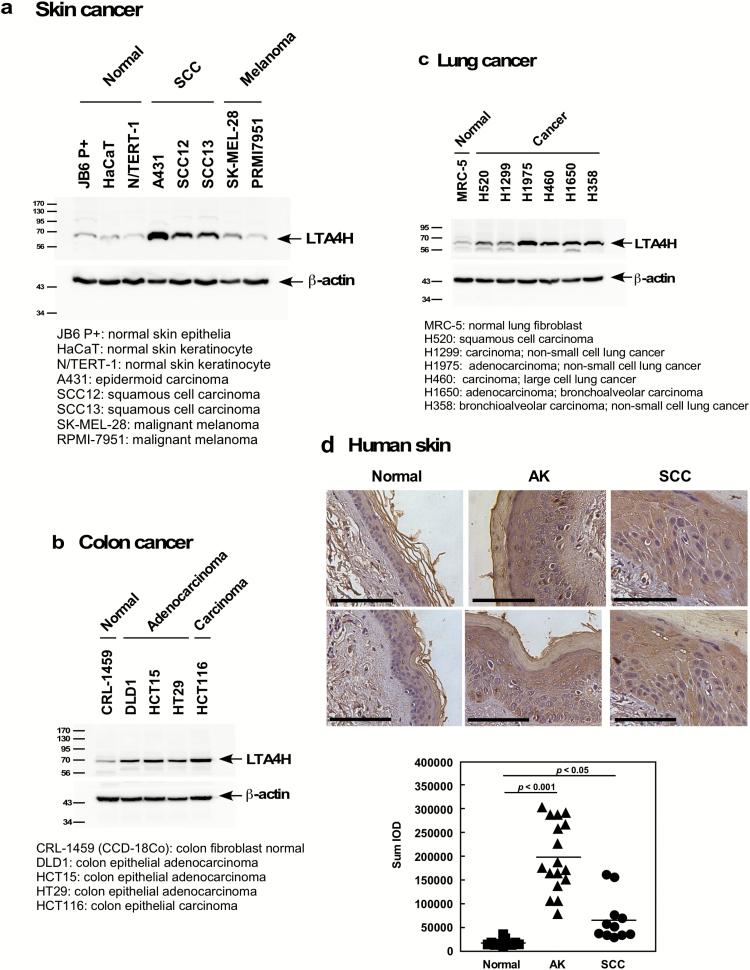
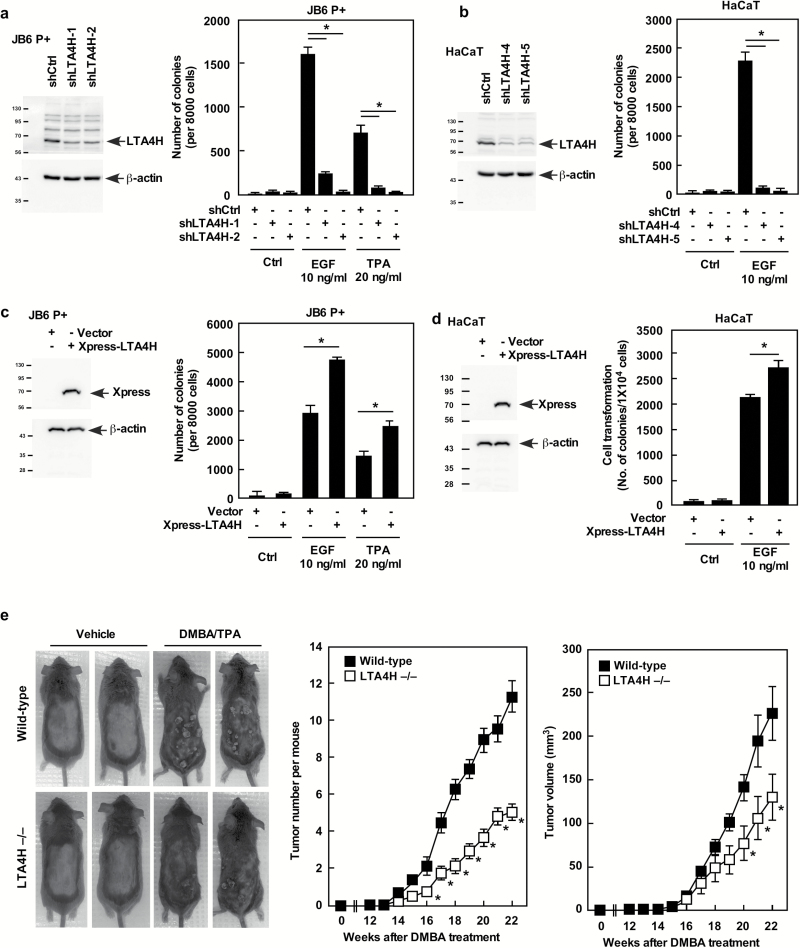
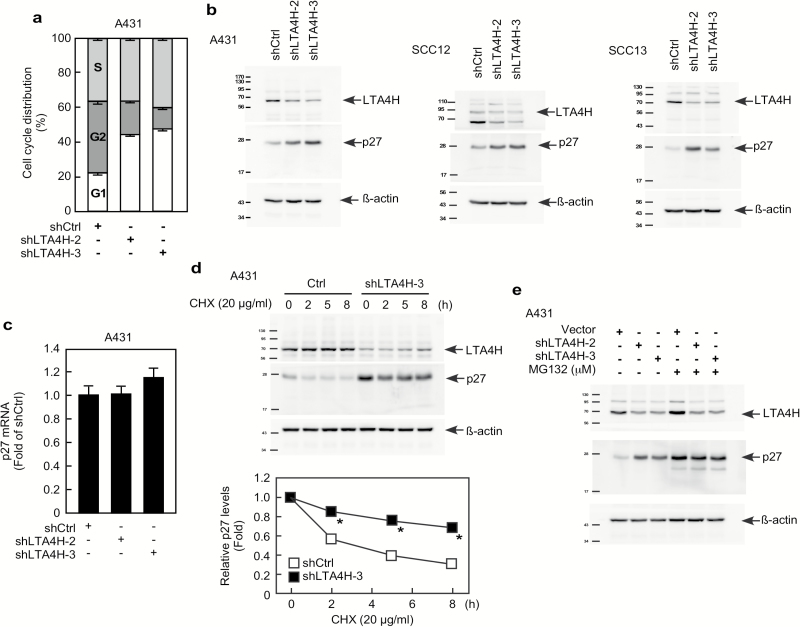
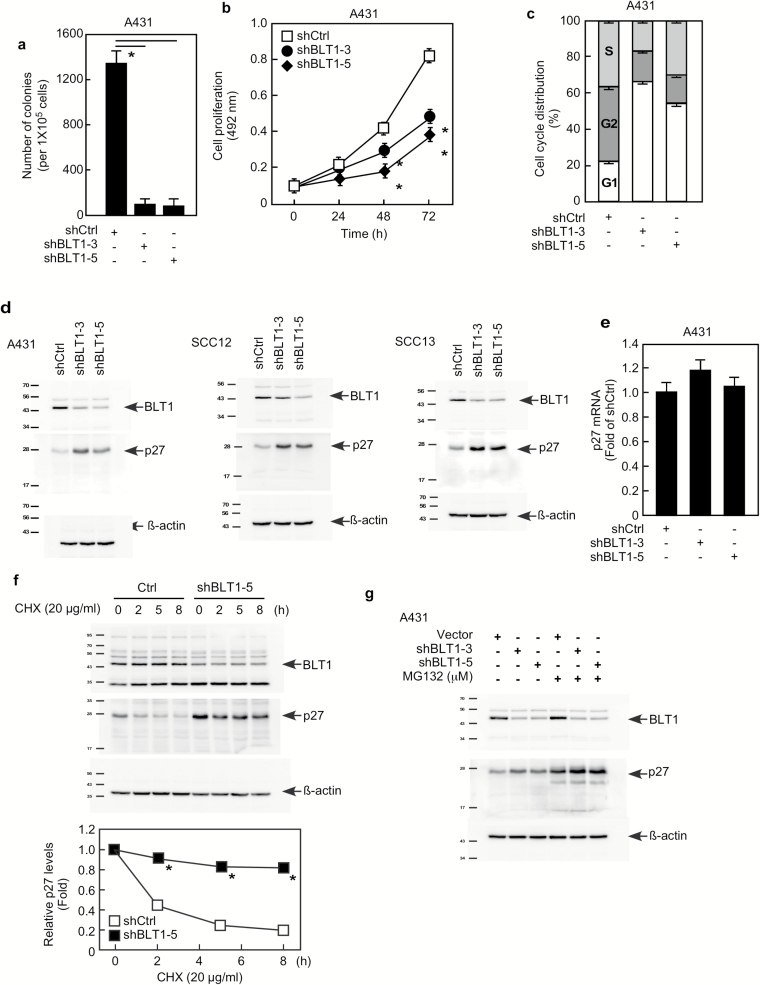
References
-
- Thunnissen M.M., et al. (2002) Crystal structures of leukotriene A4 hydrolase in complex with captopril and two competitive tight-binding inhibitors. FASEB J., 16, 1648–1650. - PubMed
-
- Andersson B., et al. (2003) Crystallization and X-ray diffraction data analysis of leukotriene A4 hydrolase from Saccharomyces cerevisiae. Acta Crystallogr. D. Biol. Crystallogr., 59, 1093–1095. - PubMed
-
- Stockley R.A., et al. (2002) The effect of augmentation therapy on bronchial inflammation in alpha1-antitrypsin deficiency. Am. J. Respir. Crit. Care Med., 165, 1494–1498. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
