

Moran's I quantifies spatio-temporal pattern formation in neural imaging data
- PMID: 28575207
- PMCID: PMC5870747
- DOI: 10.1093/bioinformatics/btx351
Moran's I quantifies spatio-temporal pattern formation in neural imaging data
Abstract
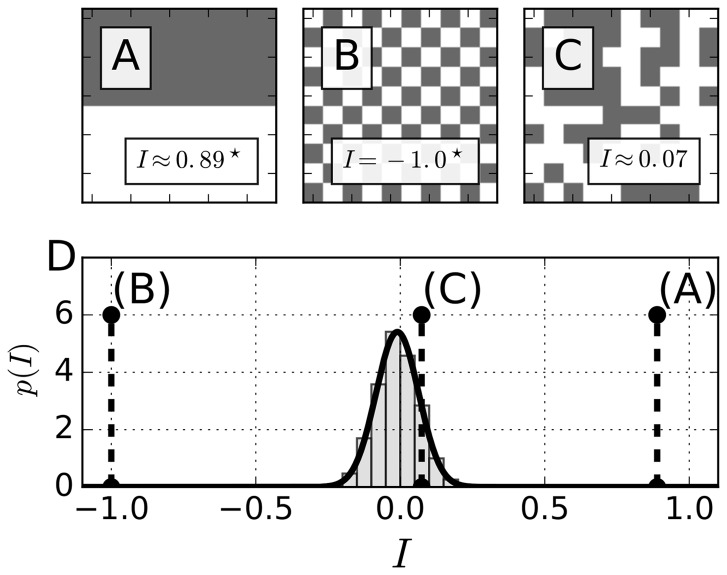
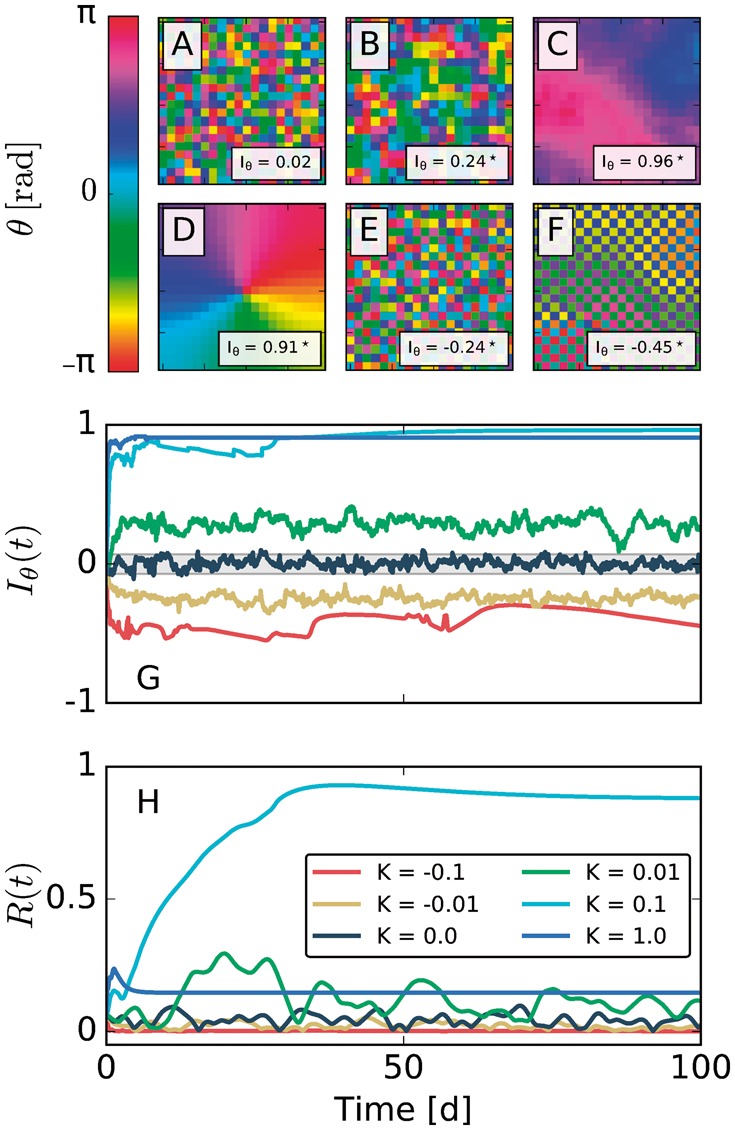
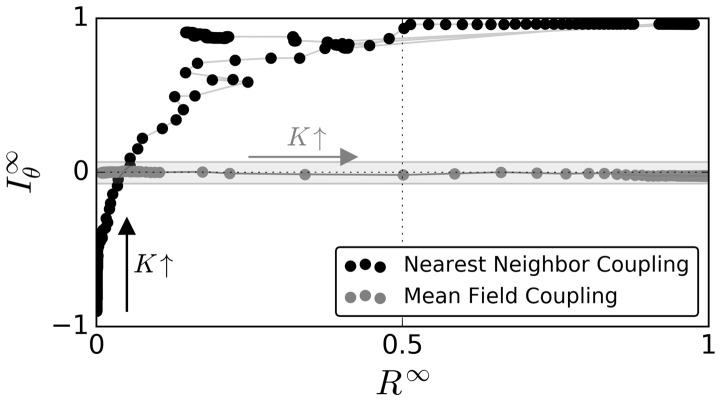
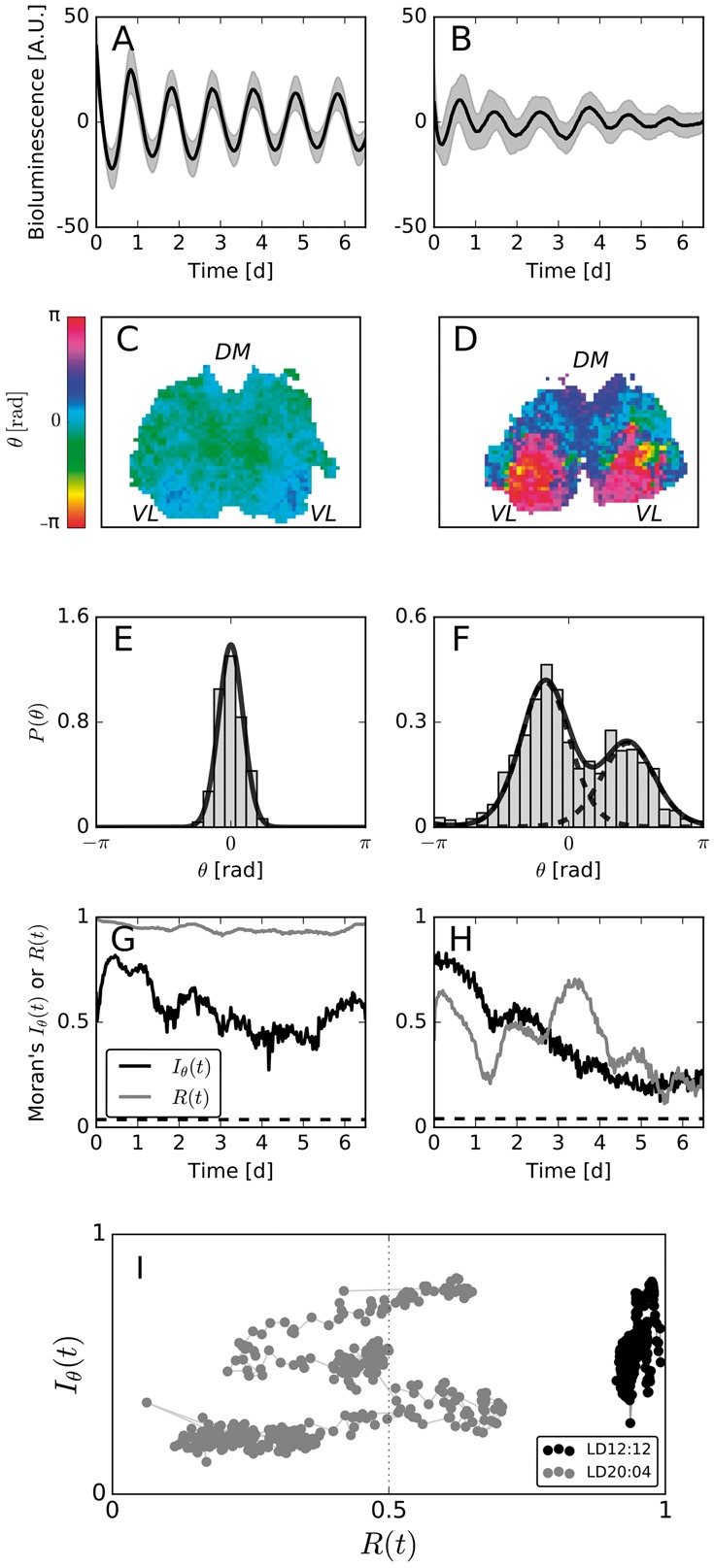
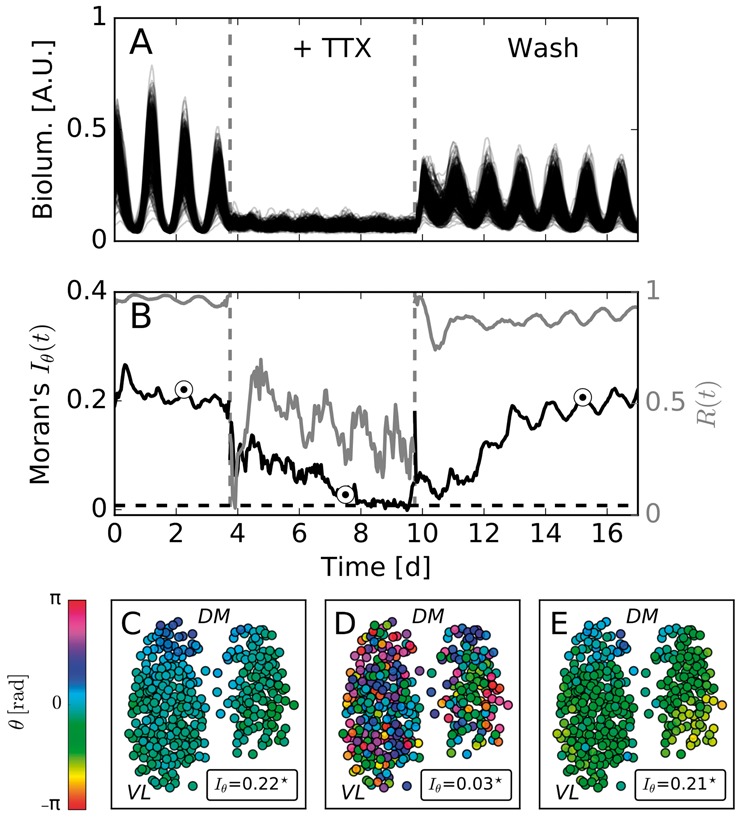
Motivation: Neural activities of the brain occur through the formation of spatio-temporal patterns. In recent years, macroscopic neural imaging techniques have produced a large body of data on these patterned activities, yet a numerical measure of spatio-temporal coherence has often been reduced to the global order parameter, which does not uncover the degree of spatial correlation. Here, we propose to use the spatial autocorrelation measure Moran's I, which can be applied to capture dynamic signatures of spatial organization. We demonstrate the application of this technique to collective cellular circadian clock activities measured in the small network of the suprachiasmatic nucleus (SCN) in the hypothalamus.
Results: We found that Moran's I is a practical quantitative measure of the degree of spatial coherence in neural imaging data. Initially developed with a geographical context in mind, Moran's I accounts for the spatial organization of any interacting units. Moran's I can be modified in accordance with the characteristic length scale of a neural activity pattern. It allows a quantification of statistical significance levels for the observed patterns. We describe the technique applied to synthetic datasets and various experimental imaging time-series from cultured SCN explants. It is demonstrated that major characteristics of the collective state can be described by Moran's I and the traditional Kuramoto order parameter R in a complementary fashion.
Availability and implementation: Python 2.7 code of illustrative examples can be found in the Supplementary Material.
Contact: christoph.schmal@charite.de or grigory.bordyugov@hu-berlin.de.
Supplementary information: Supplementary data are available at Bioinformatics online.
© The Author(s) 2017. Published by Oxford University Press.
Figures
References
-
- Acebrón J.A. et al. (2005) The Kuramoto model: a simple paradigm for synchronization phenomena. Rev. Mod. Phys., 77, 137–185.
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
