

European contribution to the study of ROS: A summary of the findings and prospects for the future from the COST action BM1203 (EU-ROS)
- PMID: 28577489
- PMCID: PMC5458069
- DOI: 10.1016/j.redox.2017.05.007
European contribution to the study of ROS: A summary of the findings and prospects for the future from the COST action BM1203 (EU-ROS)
Erratum in
-
Corrigendum to "European contribution to the study of ROS: A summary of the findings and prospects for the future from the COST action BM1203 (EU-ROS)" [Redox Biol. 13 (2017) 94-162].Redox Biol. 2018 Apr;14:694-696. doi: 10.1016/j.redox.2017.10.001. Epub 2017 Oct 26. Redox Biol. 2018. PMID: 29107648 Free PMC article. No abstract available.
Abstract
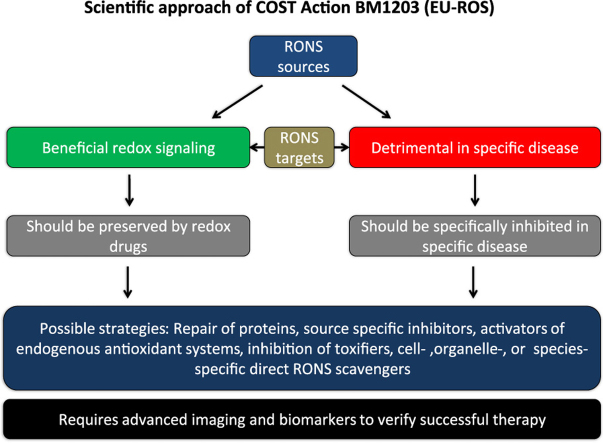
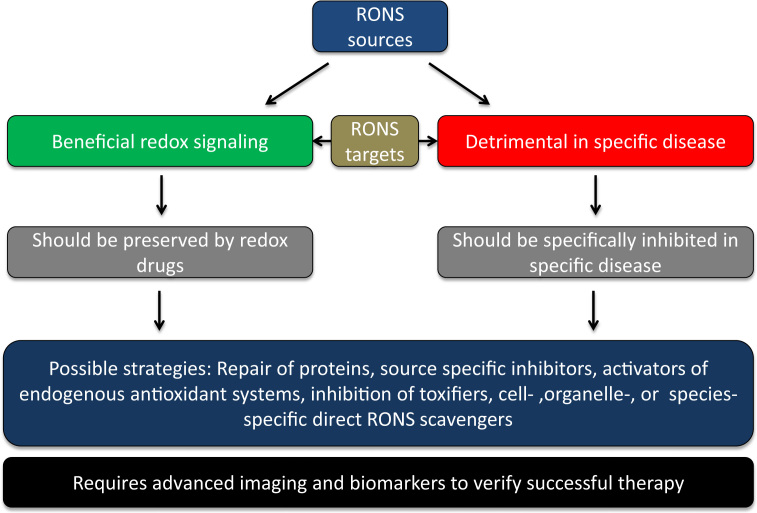
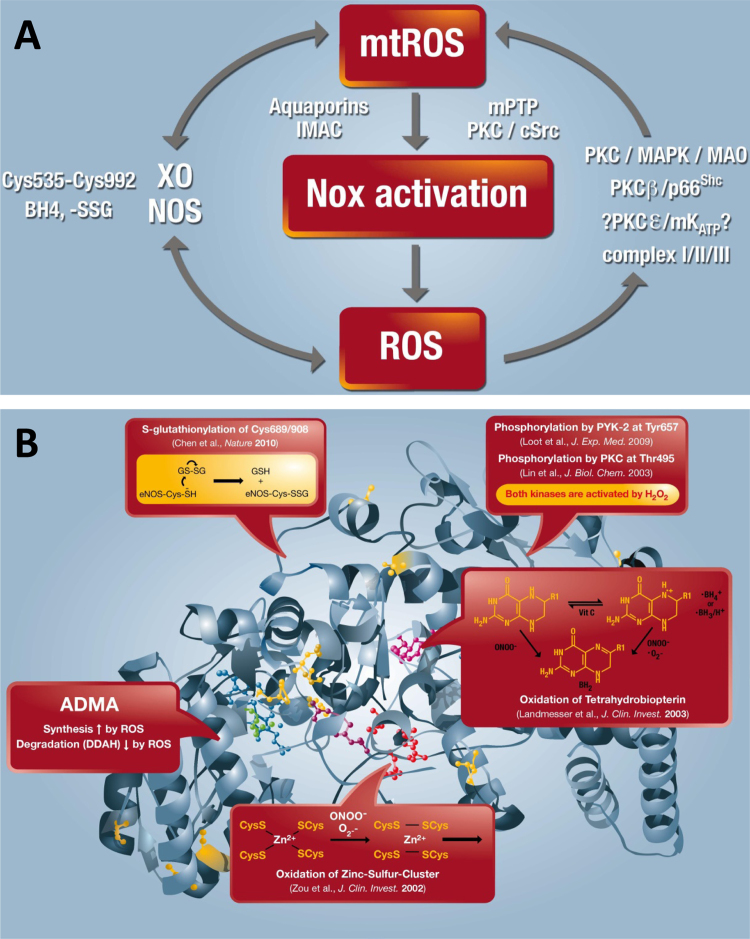
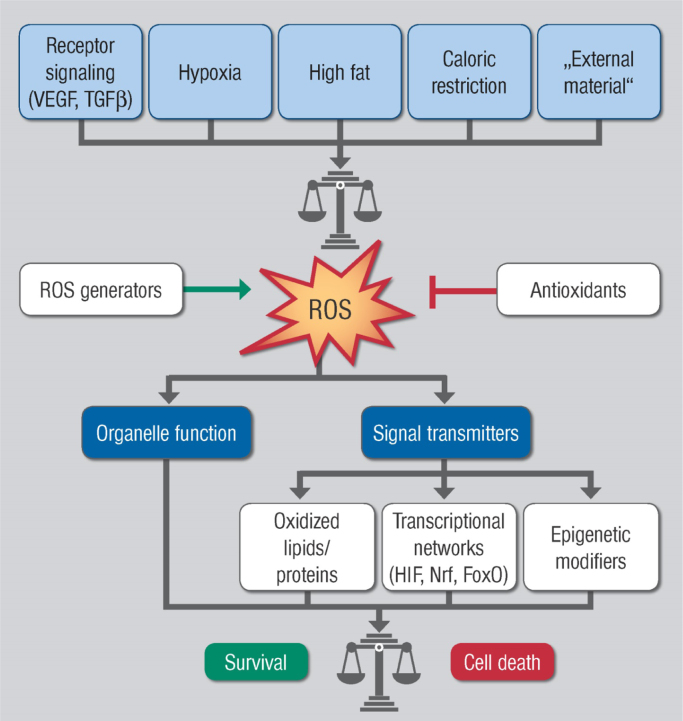
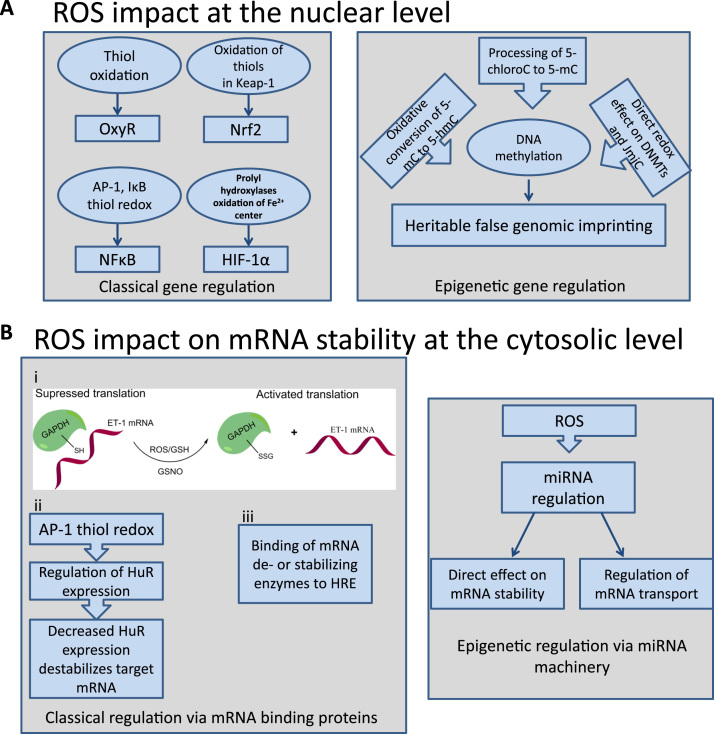
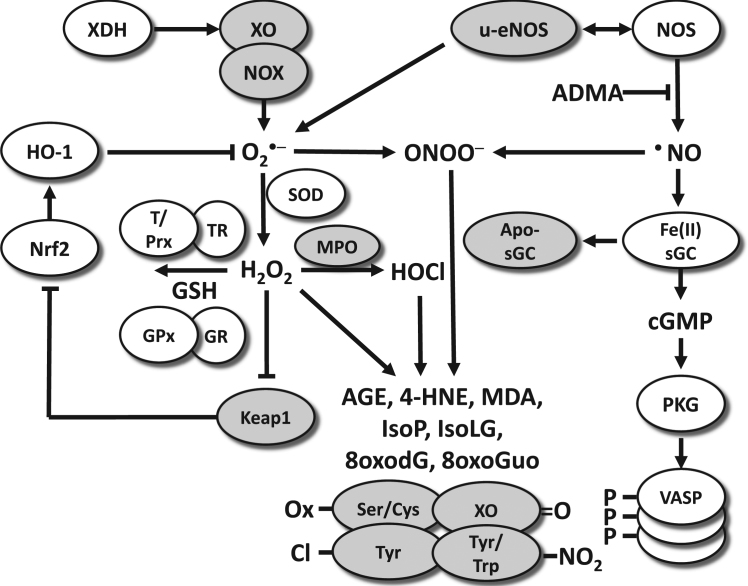
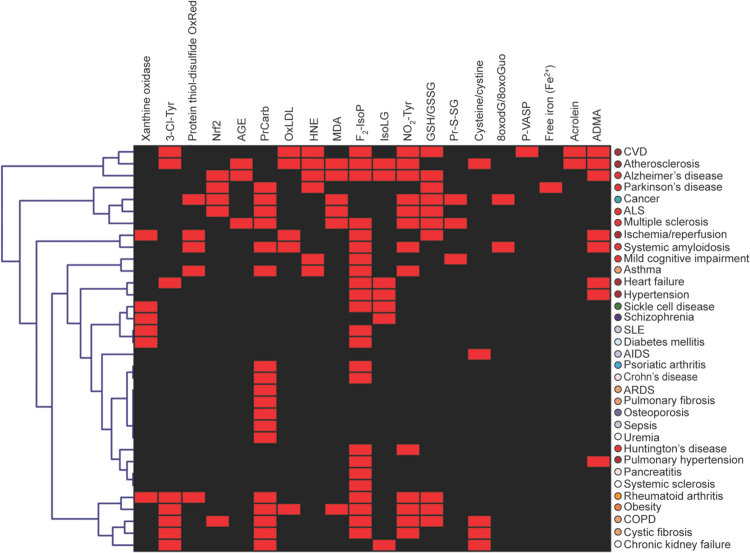
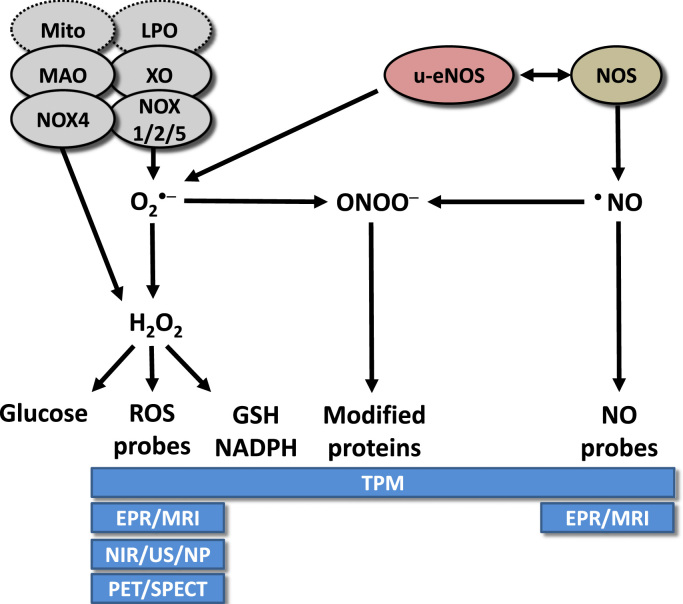
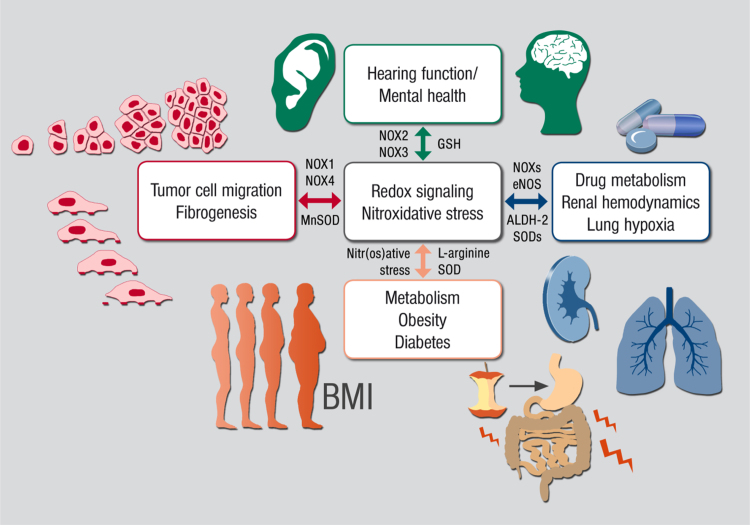
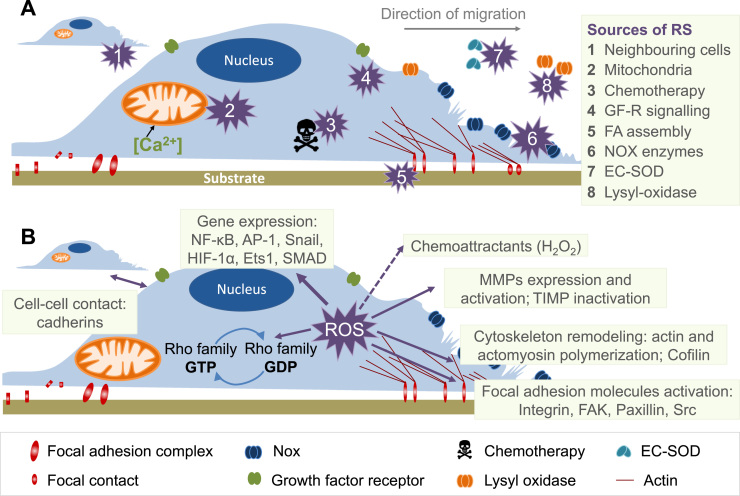
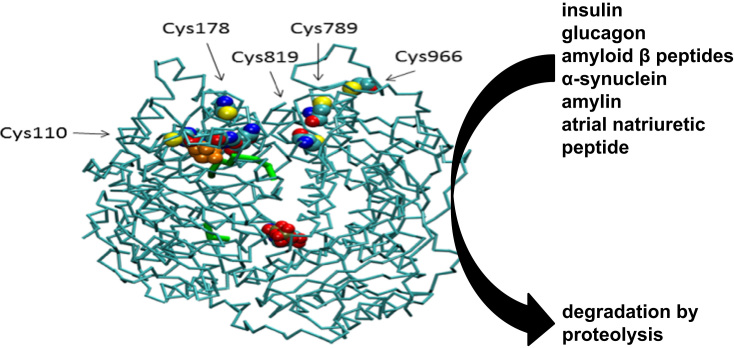
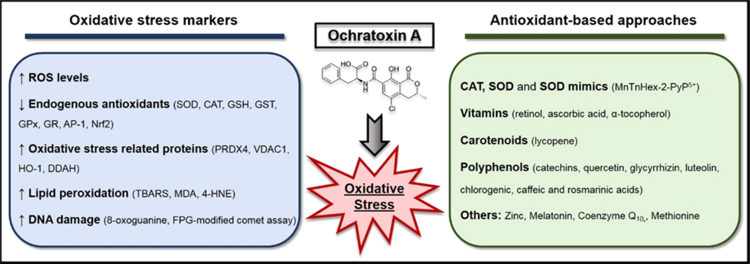
The European Cooperation in Science and Technology (COST) provides an ideal framework to establish multi-disciplinary research networks. COST Action BM1203 (EU-ROS) represents a consortium of researchers from different disciplines who are dedicated to providing new insights and tools for better understanding redox biology and medicine and, in the long run, to finding new therapeutic strategies to target dysregulated redox processes in various diseases. This report highlights the major achievements of EU-ROS as well as research updates and new perspectives arising from its members. The EU-ROS consortium comprised more than 140 active members who worked together for four years on the topics briefly described below. The formation of reactive oxygen and nitrogen species (RONS) is an established hallmark of our aerobic environment and metabolism but RONS also act as messengers via redox regulation of essential cellular processes. The fact that many diseases have been found to be associated with oxidative stress established the theory of oxidative stress as a trigger of diseases that can be corrected by antioxidant therapy. However, while experimental studies support this thesis, clinical studies still generate controversial results, due to complex pathophysiology of oxidative stress in humans. For future improvement of antioxidant therapy and better understanding of redox-associated disease progression detailed knowledge on the sources and targets of RONS formation and discrimination of their detrimental or beneficial roles is required. In order to advance this important area of biology and medicine, highly synergistic approaches combining a variety of diverse and contrasting disciplines are needed.
Keywords: Antioxidants; Oxidative stress; Reactive nitrogen species; Reactive oxygen species; Redox signaling; Redox therapeutics.
Copyright © 2017 The Authors. Published by Elsevier B.V. All rights reserved.
Figures
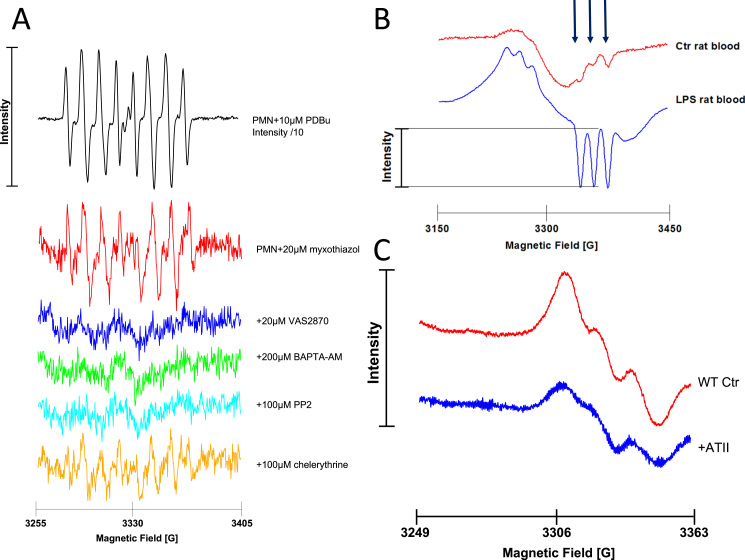
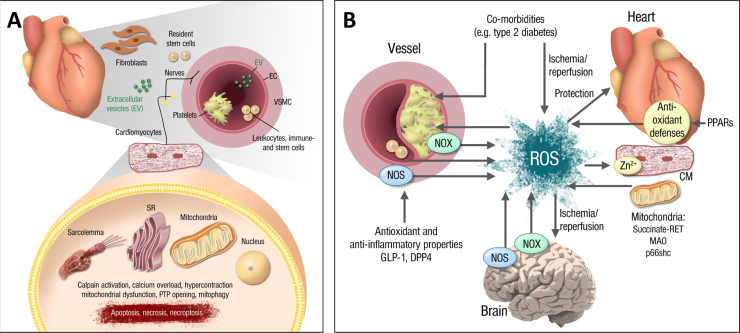
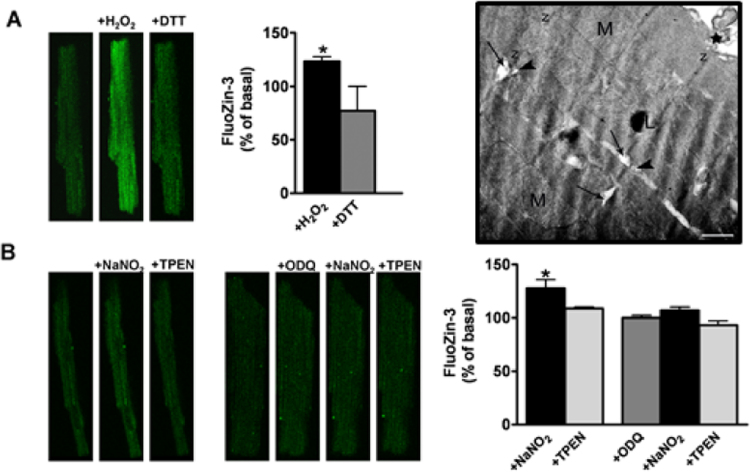
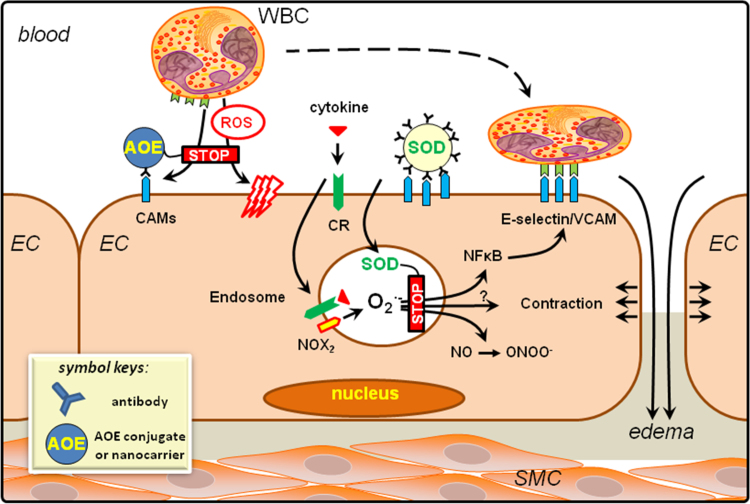
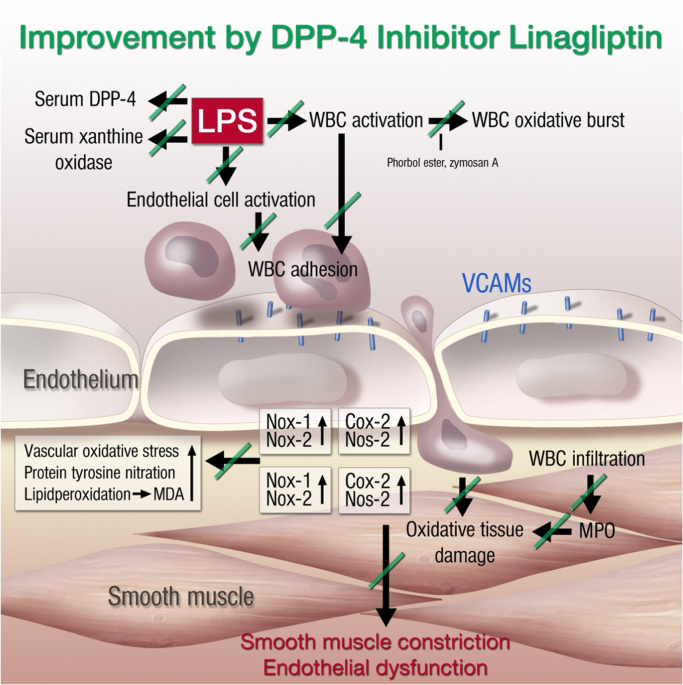
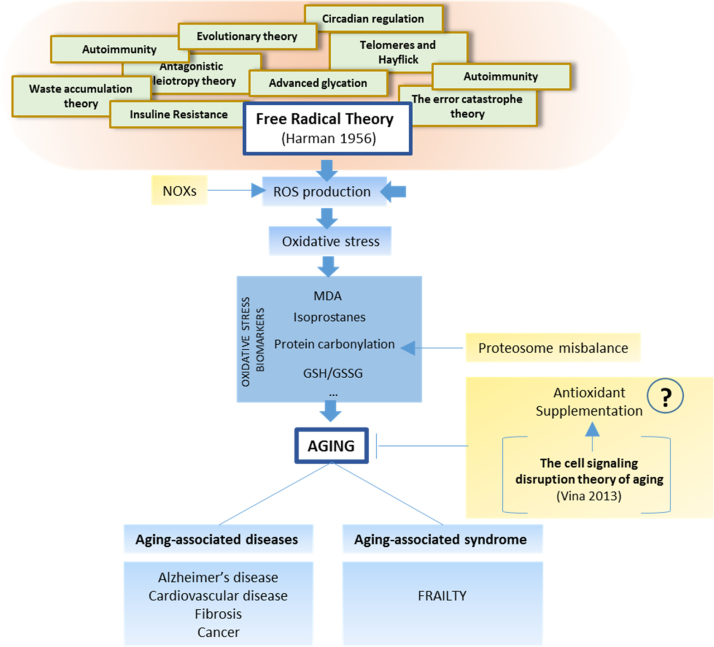
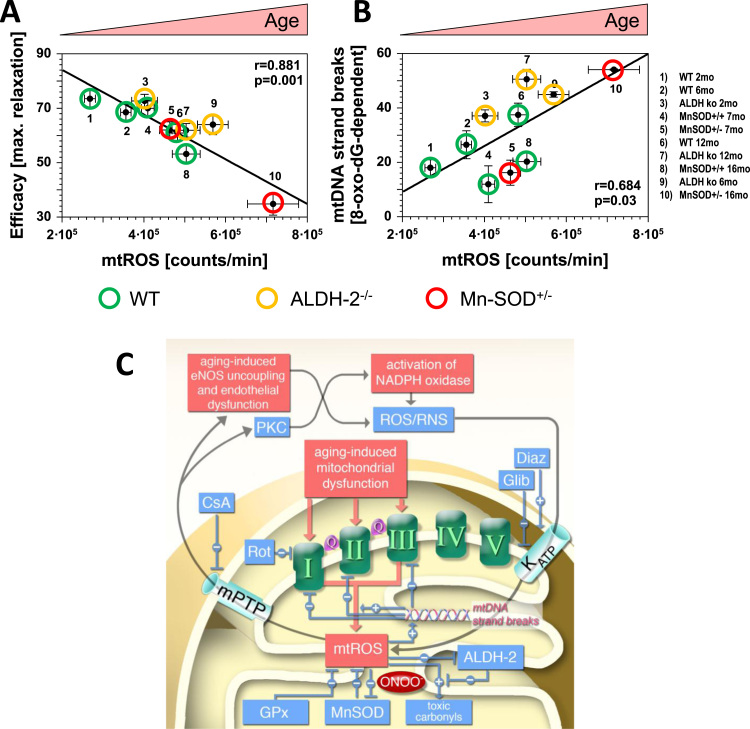
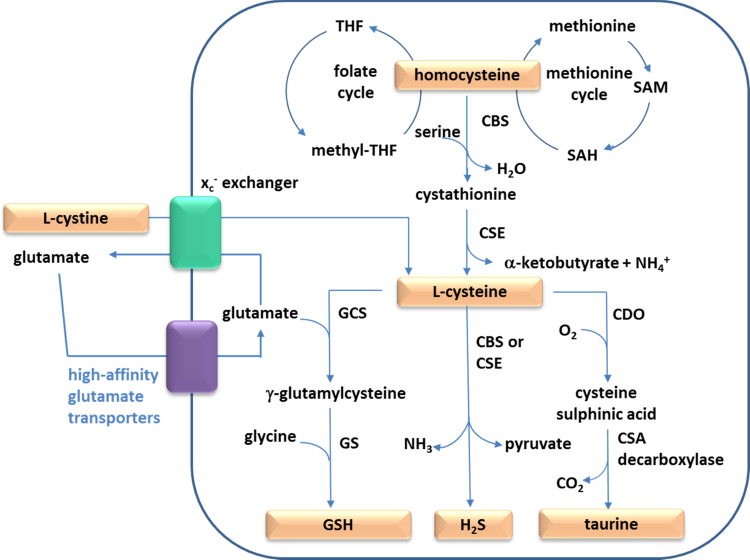
References
-
- Virtual Collection, Emerging concepts in redox biology and oxidative stress, Redox Biol. (18 articles plus editorial), in: Santiago Lamas, Fabio Di Lisa, Andreas Daiber (eds.). 〈https://www.journals.elsevier.com/redox-biology/virtual-collections/eme.... - PMC - PubMed
-
- Forum Issue, Redox medicine, Antioxid. Redox Signal. (9 articles), in: Harald H.H.W. Schmidt, Fabio Di Lisa (eds.) 〈http://online.liebertpub.com/toc/ars/23/14〉.
-
- Themed issue, Redox biology and oxidative stress in health and disease, Br. J. Pharmacol. (16 articles), in: Peter Ferdinandy and Andreas Daiber (eds.). 〈http://onlinelibrary.wiley.com/journal/10.1111/(ISSN)1476–5381/homepage/....
-
- Augusto O., Miyamoto S. Oxygen radicals and related species. In: Pantopoulos K., Schipper H.M., editors. Principles of Free Radical Biomedicine. Nova Science Publishers, Inc; 2011.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
