

Sphingosine 1-phosphate receptor 3 and RhoA signaling mediate inflammatory gene expression in astrocytes
- PMID: 28577576
- PMCID: PMC5455202
- DOI: 10.1186/s12974-017-0882-x
Sphingosine 1-phosphate receptor 3 and RhoA signaling mediate inflammatory gene expression in astrocytes
Abstract
Background: Sphingosine 1-phosphate (S1P) signals through G protein-coupled receptors to elicit a wide range of cellular responses. In CNS injury and disease, the blood-brain barrier is compromised, causing leakage of S1P from blood into the brain. S1P can also be locally generated through the enzyme sphingosine kinase-1 (Sphk1). Our previous studies demonstrated that S1P activates inflammation in murine astrocytes. The S1P1 receptor subtype has been most associated with CNS disease, particularly multiple sclerosis. S1P3 is most highly expressed and upregulated on astrocytes, however, thus we explored the involvement of this receptor in inflammatory astrocytic responses.
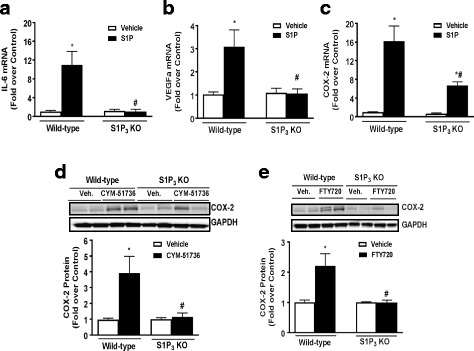
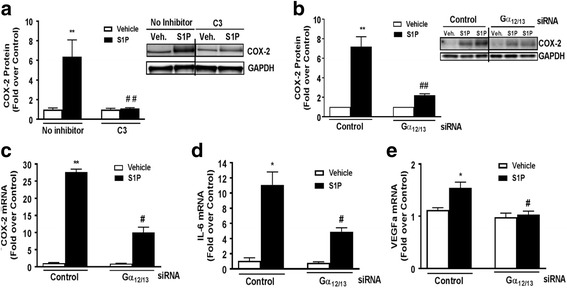
Methods: Astrocytes isolated from wild-type (WT) or S1P3 knockout (KO) mice were treated with S1P3 selective drugs or transfected with short interfering RNA to determine which receptor subtypes mediate S1P-stimulated inflammatory responses. Interleukin-6 (IL-6), and vascular endothelial growth factor A (VEGFa) messenger RNA (mRNA) and cyclooxygenase-2 (COX-2) mRNA and protein were assessed by q-PCR and Western blotting. Activation of RhoA was measured using SRE.L luciferase and RhoA implicated in S1P signaling by knockdown of Gα12/13 proteins or by inhibiting RhoA activation with C3 exoenzyme. Inflammation was simulated by in vitro scratch injury of cultured astrocytes.
Results: S1P3 was highly expressed in astrocytes and further upregulated in response to simulated inflammation. Studies using S1P3 knockdown and S1P3 KO astrocytes demonstrated that S1P3 mediates activation of RhoA and induction of COX-2, IL-6, and VEGFa mRNA, with some contribution from S1P2. S1P induces expression of all of these genes through coupling to the Gα12/13 proteins which activate RhoA. Studies using S1P3 selective agonists/antagonists as well as Fingolimod (FTY720) confirmed that stimulation of S1P3 induces COX-2 expression in astrocytes. Simulated inflammation increased expression of Sphk1 and consequently activated S1P3, demonstrating an autocrine pathway through which S1P is formed and released from astrocytes to regulate COX-2 expression.
Conclusions: S1P3, through its ability to activate RhoA and its upregulation in astrocytes, plays a unique role in inducing inflammatory responses and should be considered as a potentially important therapeutic target for CNS disease progression.
Keywords: Astrocytes; Central nervous system; Inflammation; RhoA; S1P; S1P3.
Figures



References
-
- Yatomi Y, Ohmori T, Rile G, Kazama F, Okamoto H, Sano T, Satoh K, Kume S, Tigyi G, Igarashi Y, Ozaki Y. Sphingosine 1-phosphate as a major bioactive lysophospholipid that is released from platelets and interacts with endothelial cells. Blood. 2000;96:3431–3438. - PubMed
-
- Yatomi Y, Igarashi Y, Yang L, Hisano N, Qi R, Asazuma N, Satoh K, Ozaki Y, Kume S. Sphingosine 1-phosphate, a bioactive sphingolipid abundantly stored in platelets, is a normal constituent of human plasma and serum. J Biochem. 1997;121:969–973. doi: 10.1093/oxfordjournals.jbchem.a021681. - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
