

Electrical synapses in mammalian CNS: Past eras, present focus and future directions
- PMID: 28577972
- PMCID: PMC5705454
- DOI: 10.1016/j.bbamem.2017.05.019
Electrical synapses in mammalian CNS: Past eras, present focus and future directions
Abstract
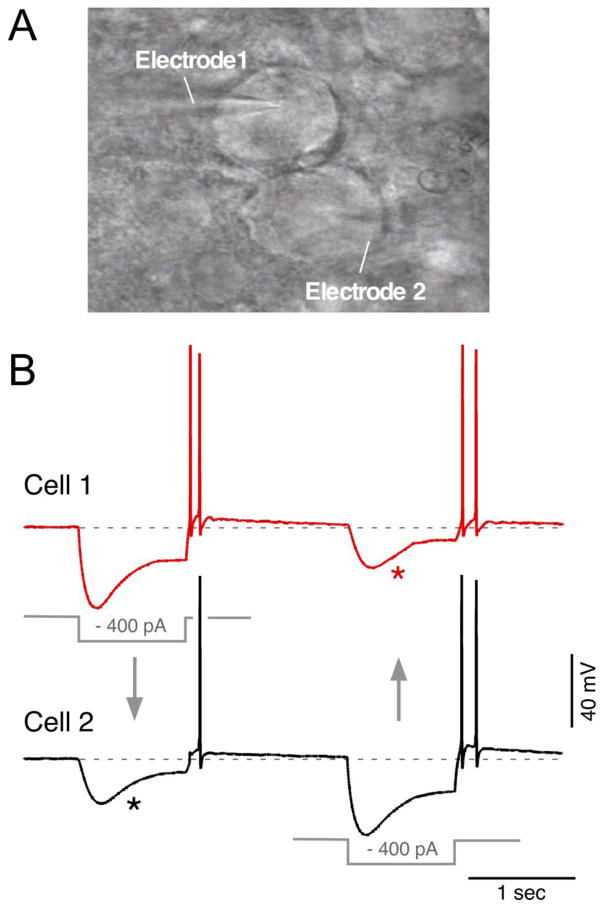
Gap junctions provide the basis for electrical synapses between neurons. Early studies in well-defined circuits in lower vertebrates laid the foundation for understanding various properties conferred by electrical synaptic transmission. Knowledge surrounding electrical synapses in mammalian systems unfolded first with evidence indicating the presence of gap junctions between neurons in various brain regions, but with little appreciation of their functional roles. Beginning at about the turn of this century, new approaches were applied to scrutinize electrical synapses, revealing the prevalence of neuronal gap junctions, the connexin protein composition of many of those junctions, and the myriad diverse neural systems in which they occur in the mammalian CNS. Subsequent progress indicated that electrical synapses constitute key elements in synaptic circuitry, govern the collective activity of ensembles of electrically coupled neurons, and in part orchestrate the synchronized neuronal network activity and rhythmic oscillations that underlie fundamental integrative processes. This article is part of a Special Issue entitled: Gap Junction Proteins edited by Jean Claude Herve.
Keywords: Cell localization; Connexins; Mixed chemical/electrical synapses; Neuronal gap junctions; Protein composition; Ultrastructural diversity.
Copyright © 2017 Elsevier B.V. All rights reserved.
Figures







References
-
- Bennett MVL, Aljure E, Nakajima Y, Pappas GD. Electrotonic junctions between teleost spinal neurons: Electrophysiology and ultrastructure. Science. 1963;141:262–264. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
