

Hsp70 facilitates trans-membrane transport of bacterial ADP-ribosylating toxins into the cytosol of mammalian cells
- PMID: 28578412
- PMCID: PMC5457432
- DOI: 10.1038/s41598-017-02882-y
Hsp70 facilitates trans-membrane transport of bacterial ADP-ribosylating toxins into the cytosol of mammalian cells
Abstract
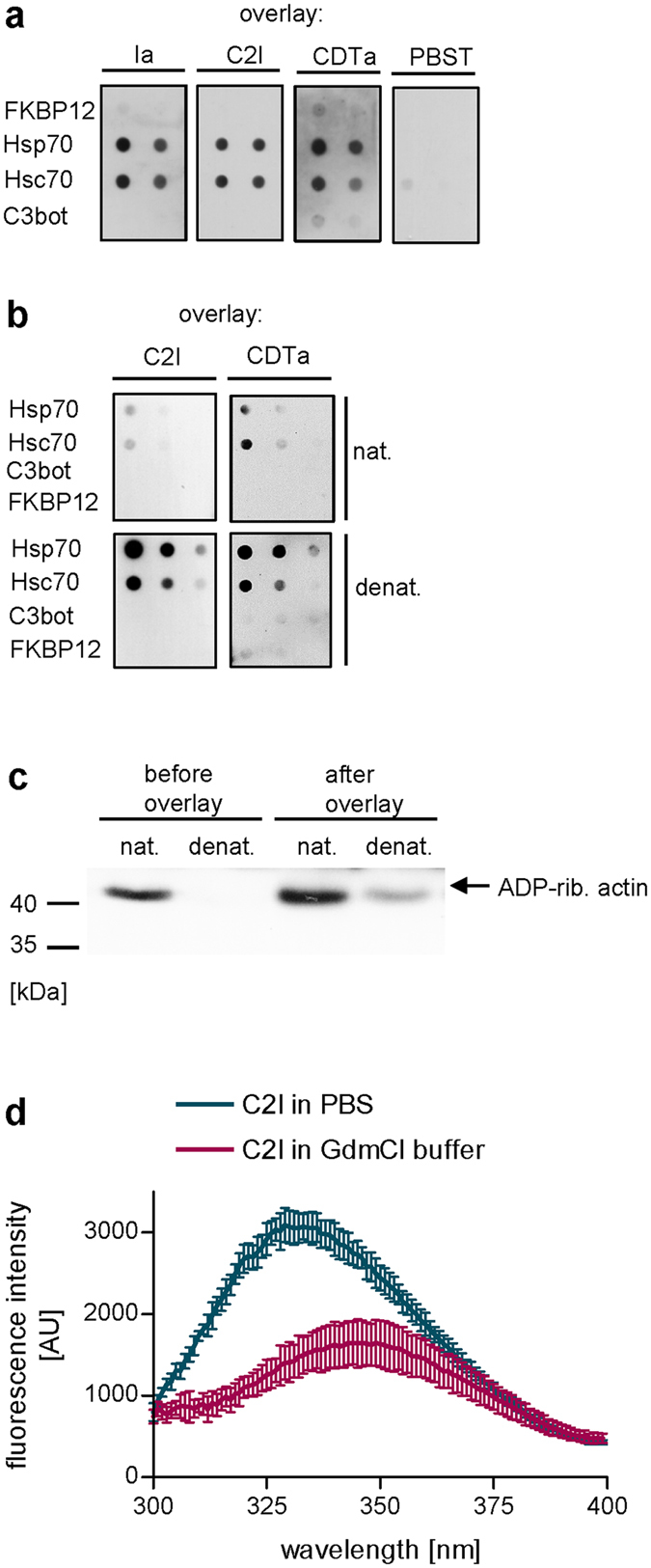
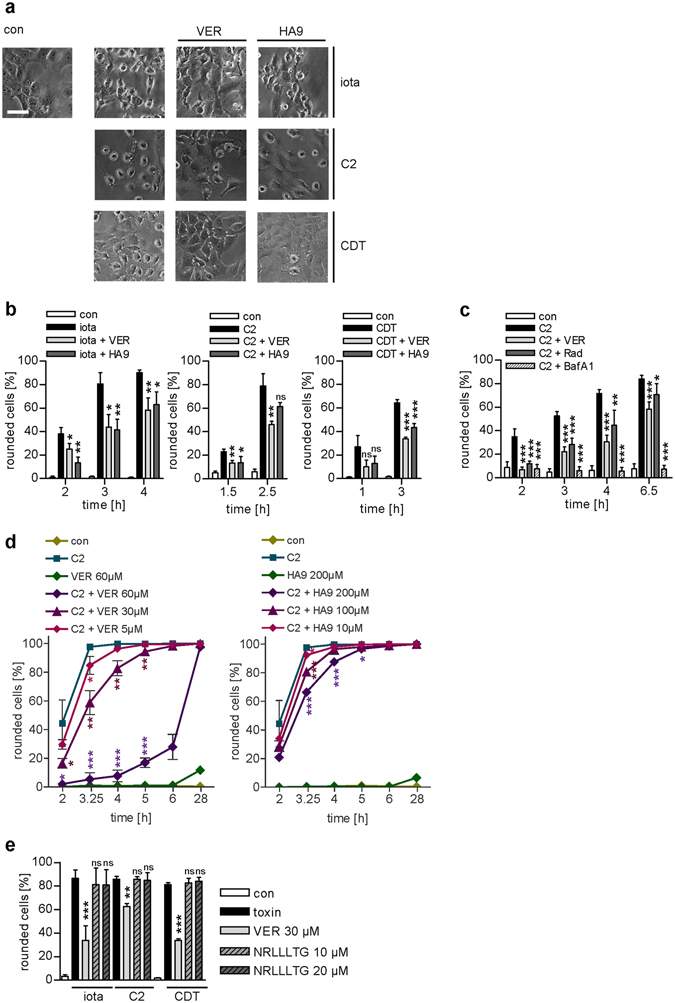
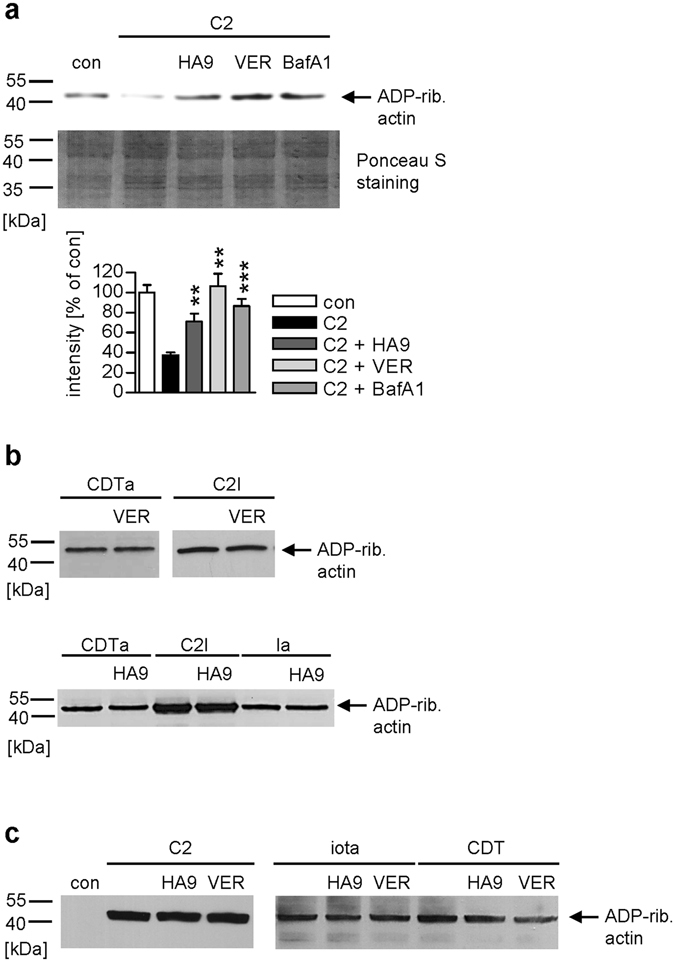
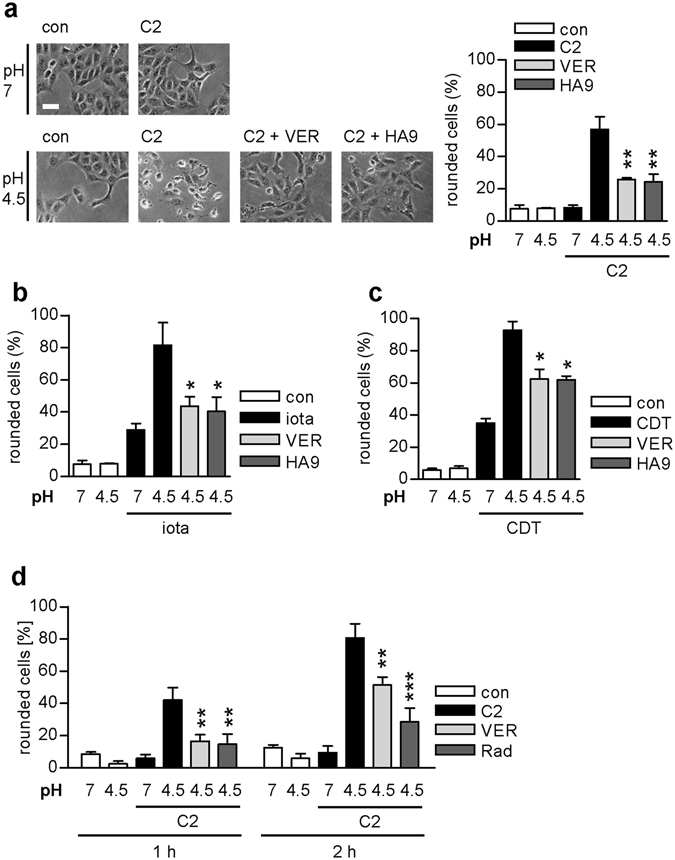
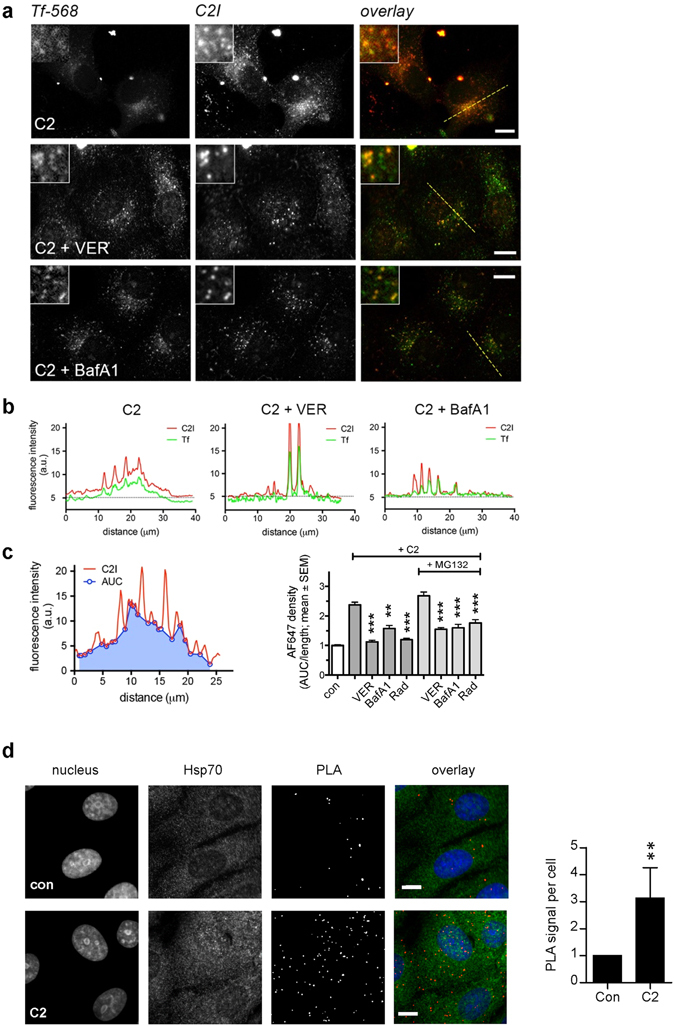
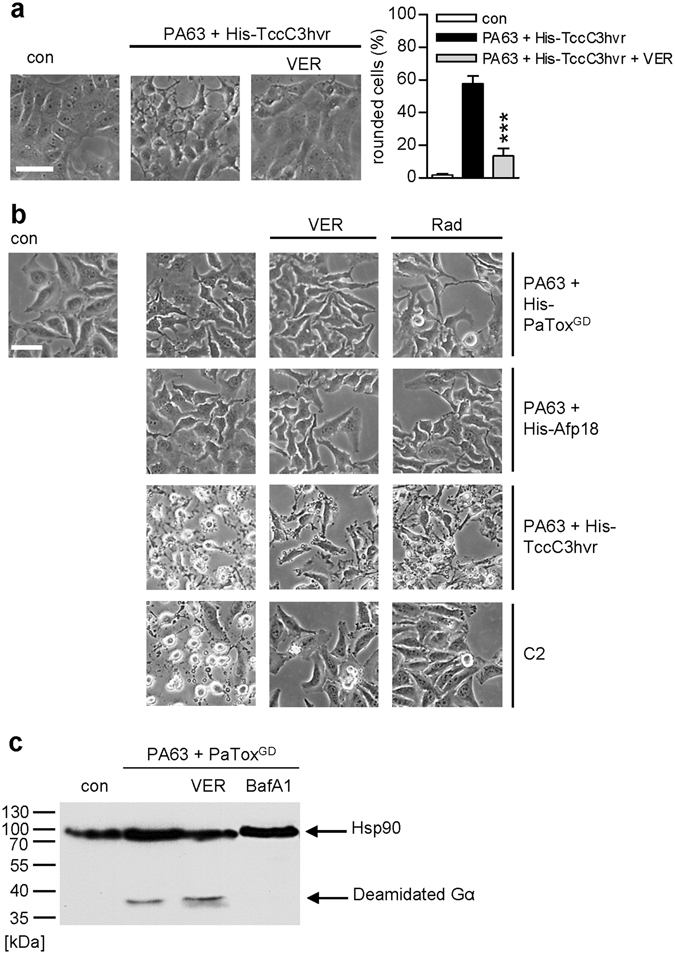
Binary enterotoxins Clostridium (C.) botulinum C2 toxin, C. perfringens iota toxin and C. difficile toxin CDT are composed of a transport (B) and a separate non-linked enzyme (A) component. Their B-components mediate endocytic uptake into mammalian cells and subsequently transport of the A-components from acidic endosomes into the cytosol, where the latter ADP-ribosylate G-actin resulting in cell rounding and cell death causing clinical symptoms. Protein folding enzymes, including Hsp90 and peptidyl-prolyl cis/trans isomerases facilitate transport of the A-components across endosomal membranes. Here, we identified Hsp70 as a novel host cell factor specifically interacting with A-components of C2, iota and CDT toxins to facilitate their transport into the cell cytosol. Pharmacological Hsp70-inhibition specifically prevented pH-dependent trans-membrane transport of A-components into the cytosol thereby protecting living cells and stem cell-derived human miniguts from intoxication. Thus, Hsp70-inhibition might lead to development of novel therapeutic strategies to treat diseases associated with bacterial ADP-ribosylating toxins.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures
Similar articles
-
EGA Protects Mammalian Cells from Clostridium difficile CDT, Clostridium perfringens Iota Toxin and Clostridium botulinum C2 Toxin.Toxins (Basel). 2016 Apr 1;8(4):101. doi: 10.3390/toxins8040101. Toxins (Basel). 2016. PMID: 27043629 Free PMC article.
-
Intoxication of mammalian cells with binary clostridial enterotoxins is inhibited by the combination of pharmacological chaperone inhibitors.Naunyn Schmiedebergs Arch Pharmacol. 2021 May;394(5):941-954. doi: 10.1007/s00210-020-02029-3. Epub 2020 Dec 7. Naunyn Schmiedebergs Arch Pharmacol. 2021. PMID: 33284399 Free PMC article.
-
Cyclophilin-facilitated membrane translocation as pharmacological target to prevent intoxication of mammalian cells by binary clostridial actin ADP-ribosylated toxins.J Mol Biol. 2015 Mar 27;427(6 Pt A):1224-38. doi: 10.1016/j.jmb.2014.07.013. Epub 2014 Jul 21. J Mol Biol. 2015. PMID: 25058685
-
Host Cell Chaperones Hsp70/Hsp90 and Peptidyl-Prolyl Cis/Trans Isomerases Are Required for the Membrane Translocation of Bacterial ADP-Ribosylating Toxins.Curr Top Microbiol Immunol. 2017;406:163-198. doi: 10.1007/82_2016_14. Curr Top Microbiol Immunol. 2017. PMID: 27197646 Review.
-
Binary actin-ADP-ribosylating toxins and their use as molecular Trojan horses for drug delivery into eukaryotic cells.Curr Med Chem. 2008;15(5):459-69. doi: 10.2174/092986708783503195. Curr Med Chem. 2008. PMID: 18289001 Review.
Cited by
-
Comparative population genomic analyses of transporters within the Asgard archaeal superphylum.PLoS One. 2021 Mar 26;16(3):e0247806. doi: 10.1371/journal.pone.0247806. eCollection 2021. PLoS One. 2021. PMID: 33770091 Free PMC article.
-
The Pore-Forming Subunit C2IIa of the Binary Clostridium botulinum C2 Toxin Reduces the Chemotactic Translocation of Human Polymorphonuclear Leukocytes.Front Pharmacol. 2022 Feb 11;13:810611. doi: 10.3389/fphar.2022.810611. eCollection 2022. Front Pharmacol. 2022. PMID: 35222028 Free PMC article.
-
Drug Inhibition of SARS-CoV-2 Replication in Human Pluripotent Stem Cell-Derived Intestinal Organoids.Cell Mol Gastroenterol Hepatol. 2021;11(4):935-948. doi: 10.1016/j.jcmgh.2020.11.003. Epub 2020 Nov 10. Cell Mol Gastroenterol Hepatol. 2021. PMID: 33186749 Free PMC article.
-
CRISPA: A Non-viral, Transient Cas9 Delivery System Based on Reengineered Anthrax Toxin.Front Pharmacol. 2021 Oct 18;12:770283. doi: 10.3389/fphar.2021.770283. eCollection 2021. Front Pharmacol. 2021. PMID: 34733166 Free PMC article.
-
Exploring the inhibitory potential of the antiarrhythmic drug amiodarone against Clostridioides difficile toxins TcdA and TcdB.Gut Microbes. 2023 Dec;15(2):2256695. doi: 10.1080/19490976.2023.2256695. Epub 2023 Sep 25. Gut Microbes. 2023. PMID: 37749884 Free PMC article.
References
-
- Kurazono H, Hosokawa M, Matsuda H, Sakaguchi G. Fluid accumulation in the ligated intestinal loop and histopathological changes of the intestinal mucosa caused by Clostridium botulinum C2 toxin in the pheasant and chicken. Res. Vet. Sci. 1987;42:349–353. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
