

Advances on the Transfer of Lipids by Lipid Transfer Proteins
- PMID: 28579073
- PMCID: PMC5486777
- DOI: 10.1016/j.tibs.2017.05.001
Advances on the Transfer of Lipids by Lipid Transfer Proteins
Abstract
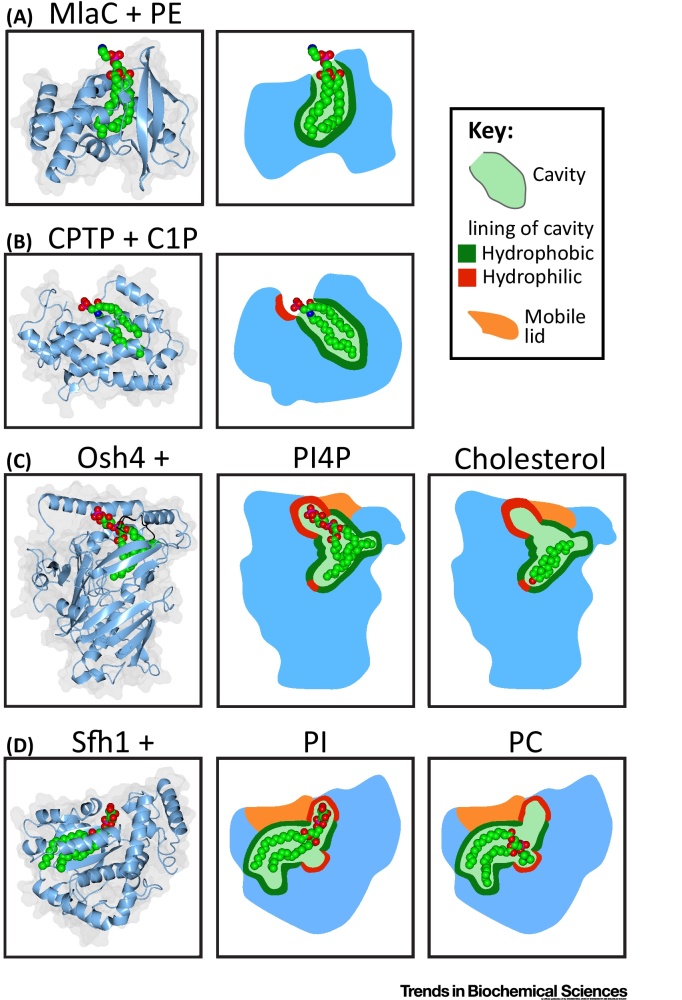
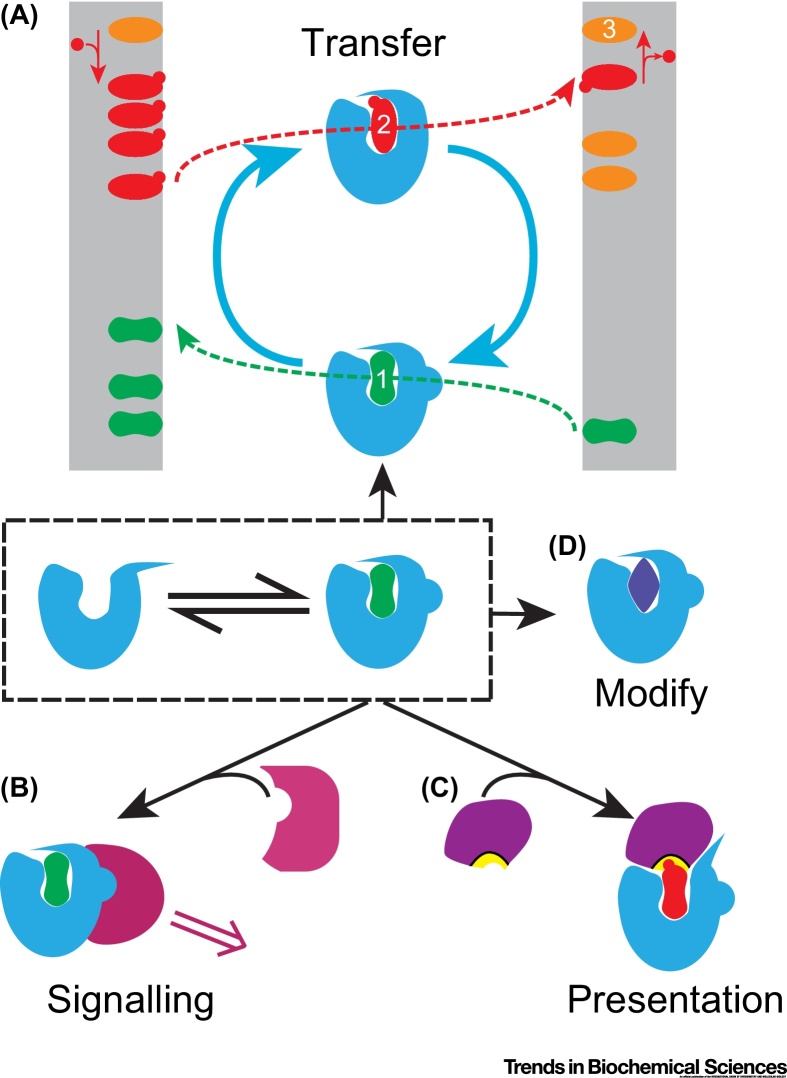
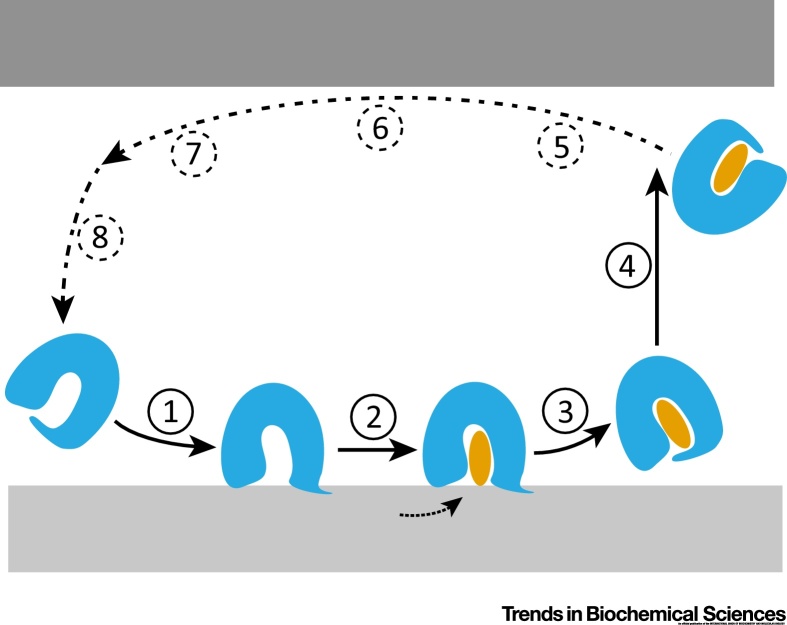
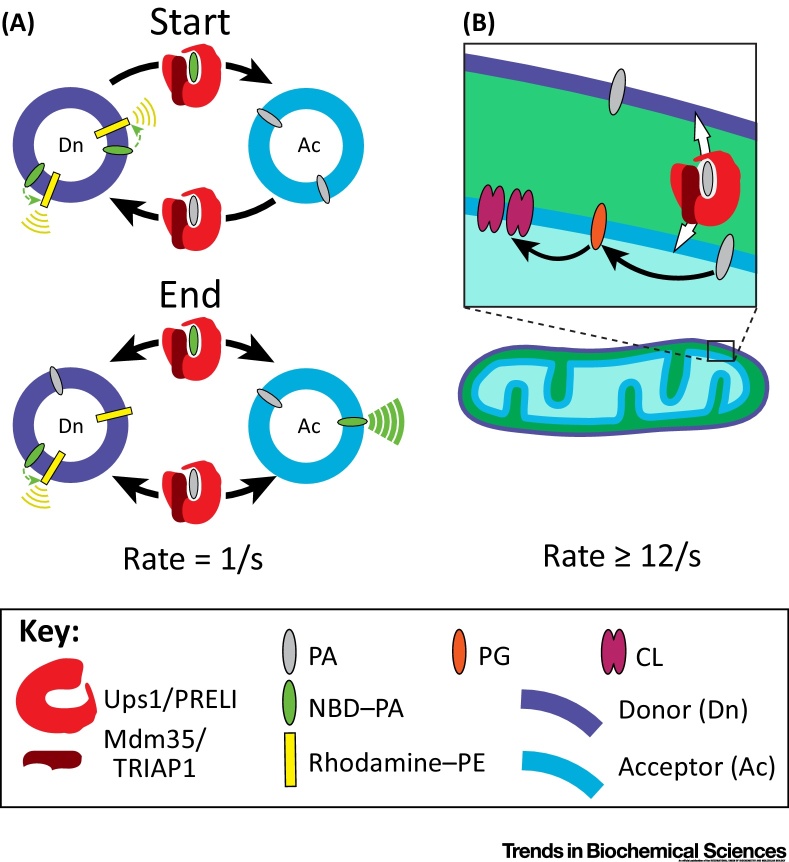
Transfer of lipid across the cytoplasm is an essential process for intracellular lipid traffic. Lipid transfer proteins (LTPs) are defined by highly controlled in vitro experiments. The functional relevance of these is supported by evidence for the same reactions inside cells. Major advances in the LTP field have come from structural bioinformatics identifying new LTPs, and from the development of countercurrent models for LTPs. However, the ultimate aim is to unite in vitro and in vivo data, and this is where much progress remains to be made. Even where in vitro and in vivo experiments align, rates of transfer tend not to match. Here we set out some of the advances that might test how LTPs work.
Keywords: lipid exchange; nonvesicular traffic; oxysterol binding protein-related proteins; tubular lipid binding proteins.
Copyright © 2017. Published by Elsevier Ltd.
Figures
References
-
- Wirtz K.W., Zilversmit D.B. Exchange of phospholipids between liver mitochondria and microsomes in vitro. J. Biol. Chem. 1968;243:3596–3602. - PubMed
-
- Daum G. Intracellular transfer of phospholipids in the yeast, Saccharomyces cerevisiae. Biochim. Biophys. Acta. 1986;878:93–101. - PubMed
-
- Pagano R.E. Lipid traffic in eukaryotic cells: mechanisms for intracellular transport and organelle-specific enrichment of lipids. Curr. Opin. Cell Biol. 1990;2:652–663. - PubMed
-
- Helmkamp G.M., Jr Phospholipid exchange between membranes. Purification of bovine brain proteins that preferentially catalyze the transfer of phosphatidylinositol. J. Biol. Chem. 1974;249:6382–6389. - PubMed
-
- Chiapparino A. The orchestra of lipid-transfer proteins at the crossroads between metabolism and signaling. Prog. Lipid Res. 2016;61:30–39. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
