

IL-17-producing γδ T cells switch migratory patterns between resting and activated states
- PMID: 28580944
- PMCID: PMC5465362
- DOI: 10.1038/ncomms15632
IL-17-producing γδ T cells switch migratory patterns between resting and activated states
Abstract
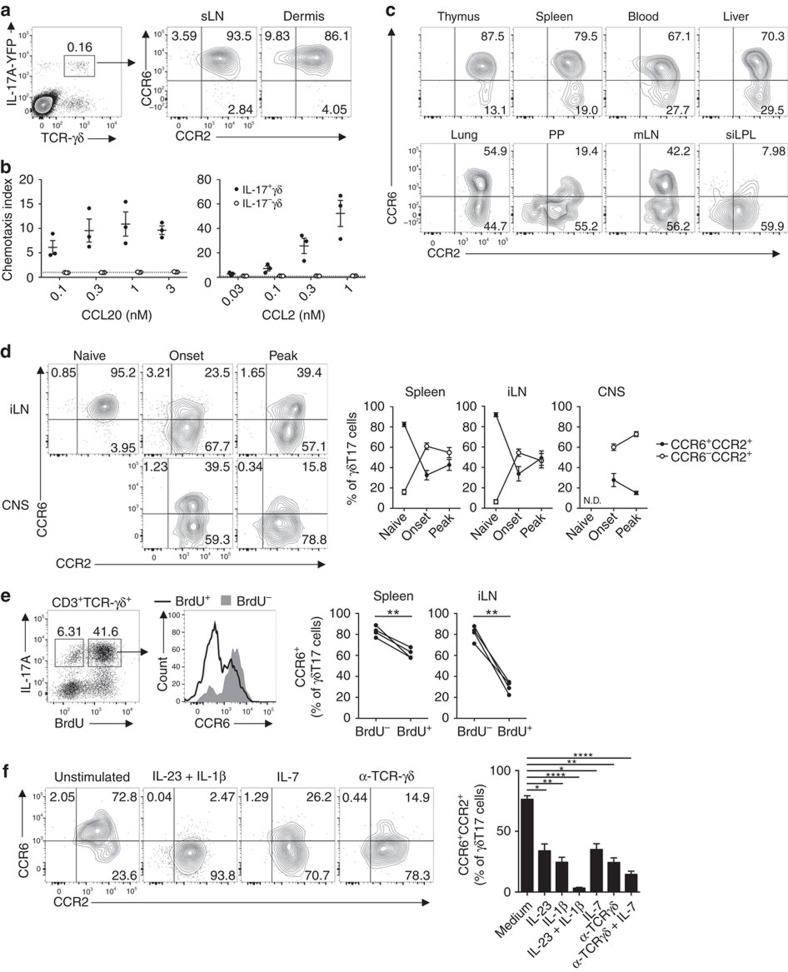
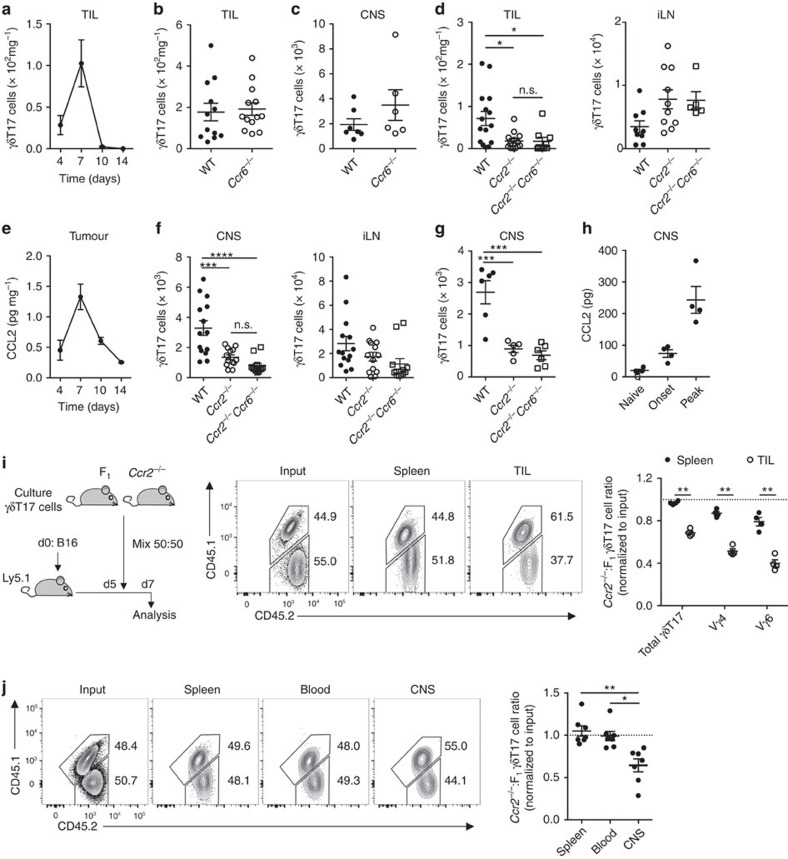
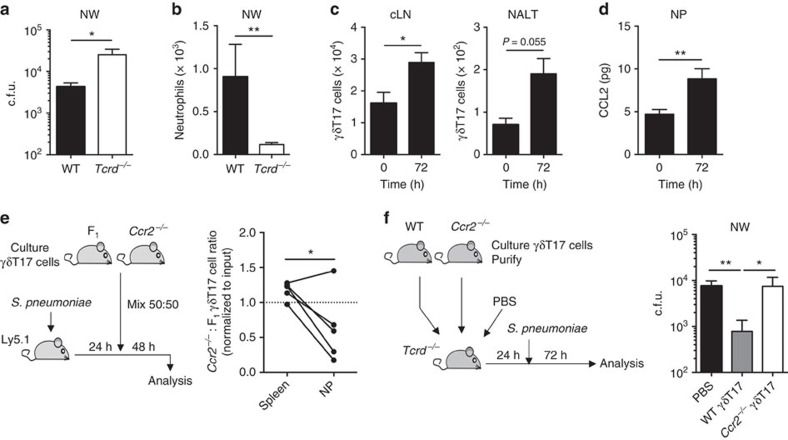
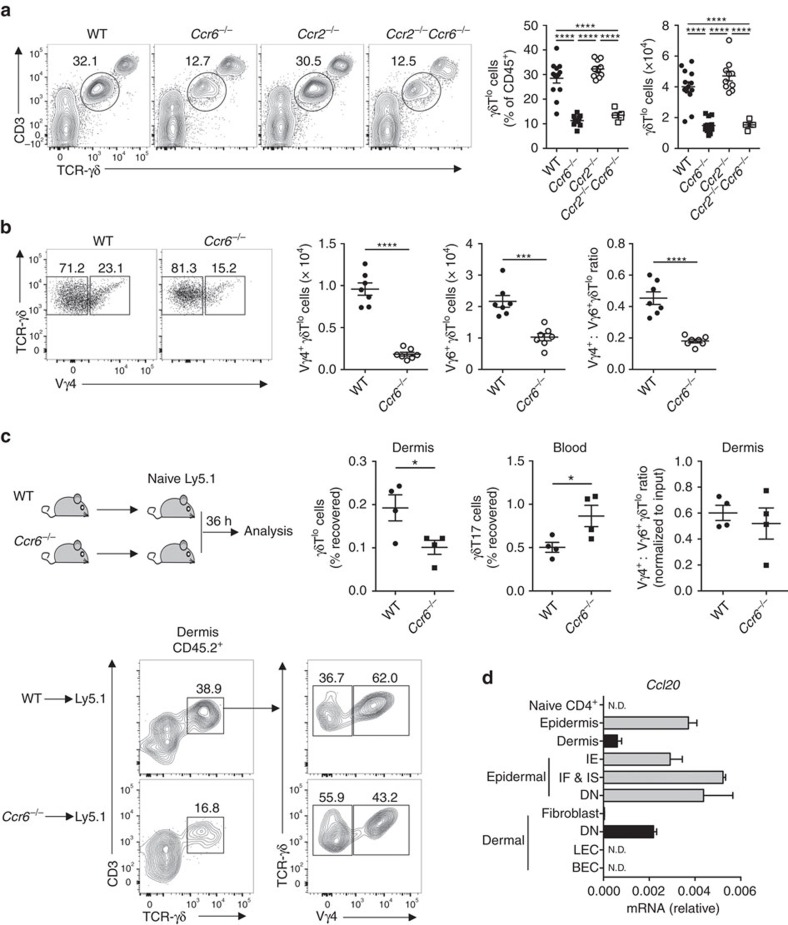
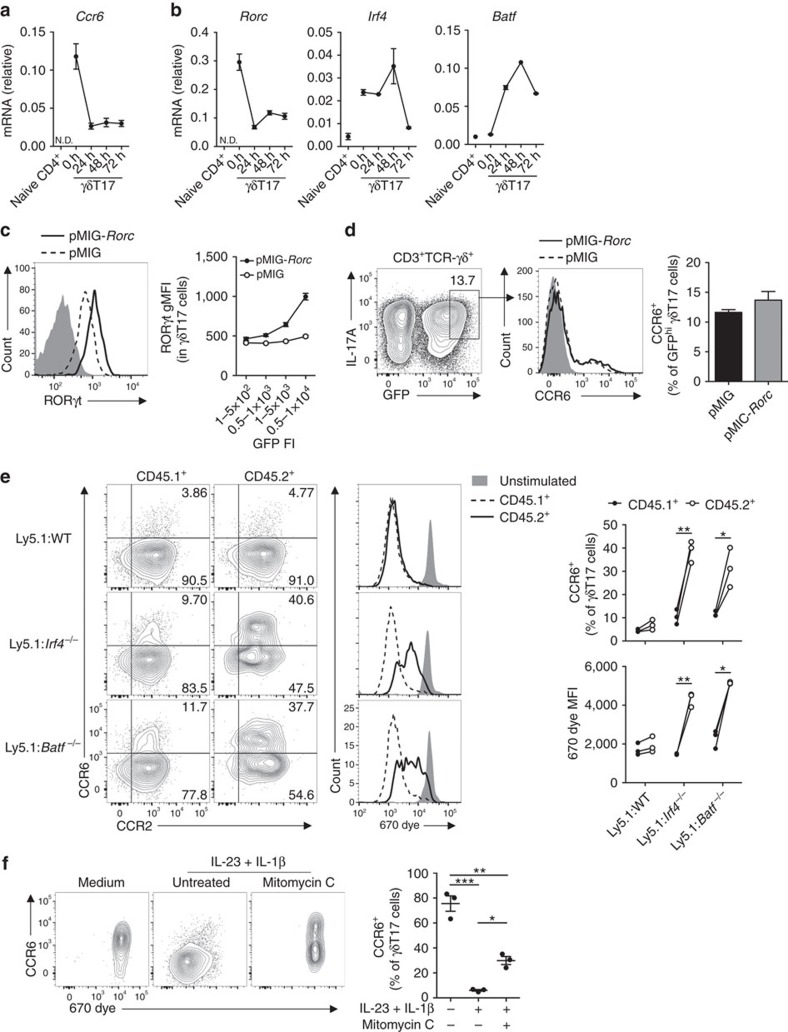
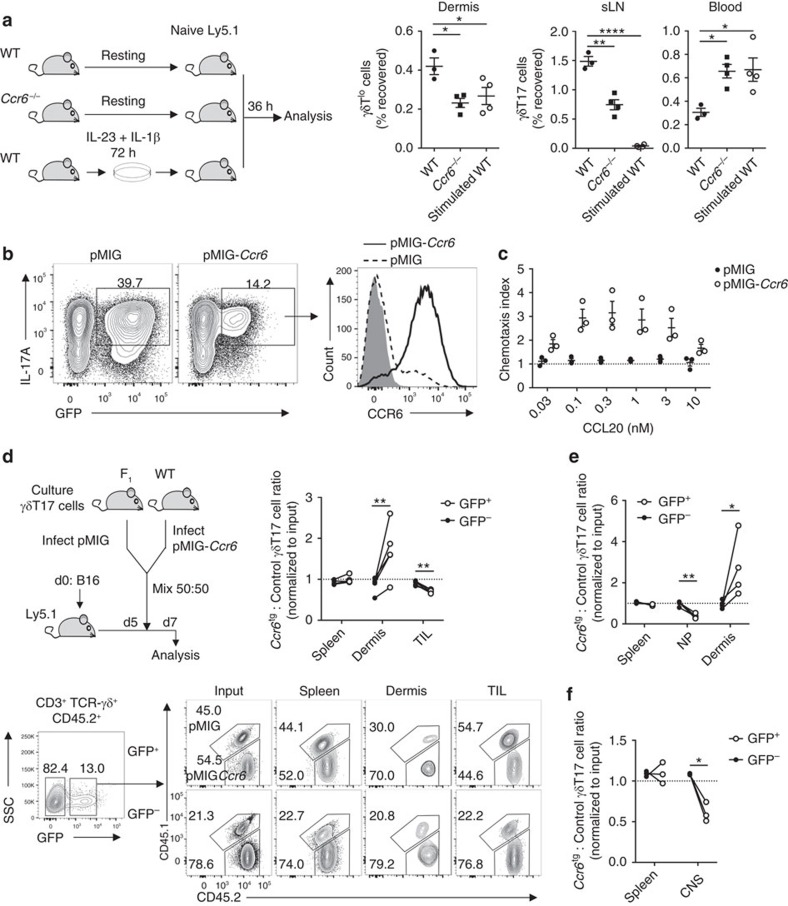
Interleukin 17-producing γδ T (γδT17) cells have unconventional trafficking characteristics, residing in mucocutaneous tissues but also homing into inflamed tissues via circulation. Despite being fundamental to γδT17-driven early protective immunity and exacerbation of autoimmunity and cancer, migratory cues controlling γδT17 cell positioning in barrier tissues and recruitment to inflammatory sites are still unclear. Here we show that γδT17 cells constitutively express chemokine receptors CCR6 and CCR2. While CCR6 recruits resting γδT17 cells to the dermis, CCR2 drives rapid γδT17 cell recruitment to inflamed tissues during autoimmunity, cancer and infection. Downregulation of CCR6 by IRF4 and BATF upon γδT17 activation is required for optimal recruitment of γδT17 cells to inflamed tissue by preventing their sequestration into uninflamed dermis. These findings establish a lymphocyte trafficking model whereby a hierarchy of homing signals is prioritized by dynamic receptor expression to drive both tissue surveillance and rapid recruitment of γδT17 cells to inflammatory lesions.
Conflict of interest statement
The authors declare no competing financial interests.
Figures
References
-
- Sutton C. E. et al. Interleukin-1 and IL-23 induce innate IL-17 production from gammadelta T cells, amplifying Th17 responses and autoimmunity. Immunity 31, 331–341 (2009). - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
