

Synergistic cooperation promotes multicellular performance and unicellular free-rider persistence
- PMID: 28580966
- PMCID: PMC5465372
- DOI: 10.1038/ncomms15707
Synergistic cooperation promotes multicellular performance and unicellular free-rider persistence
Abstract
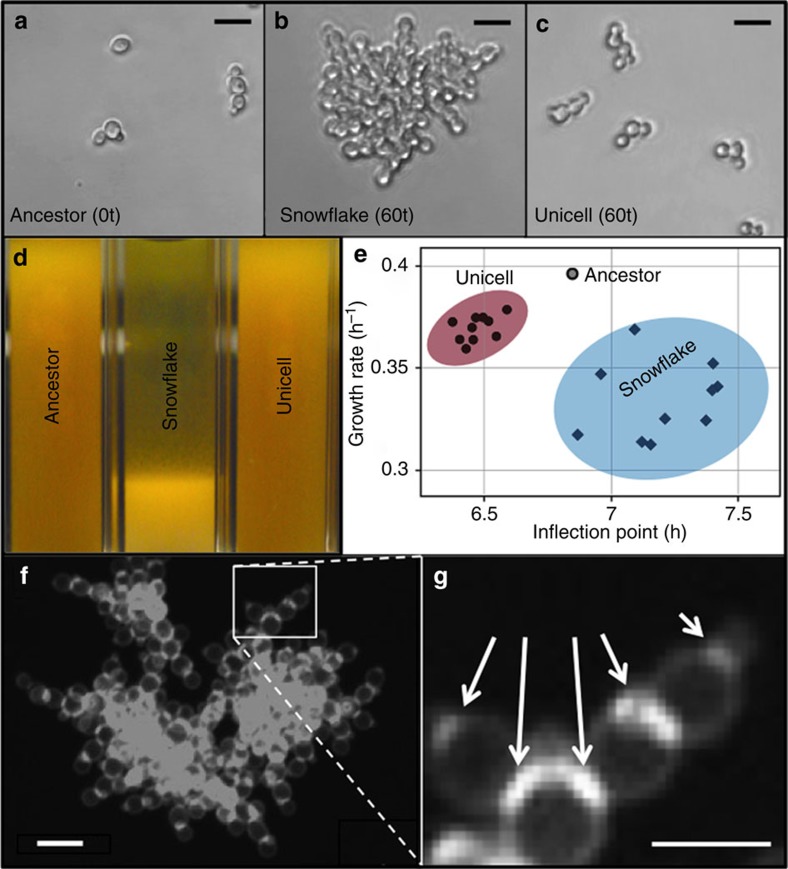
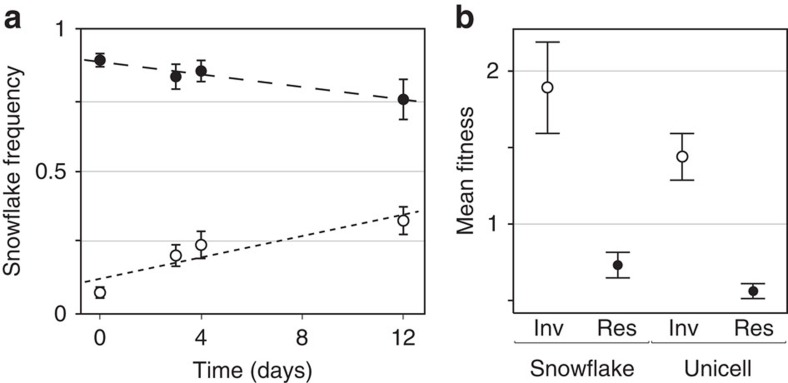
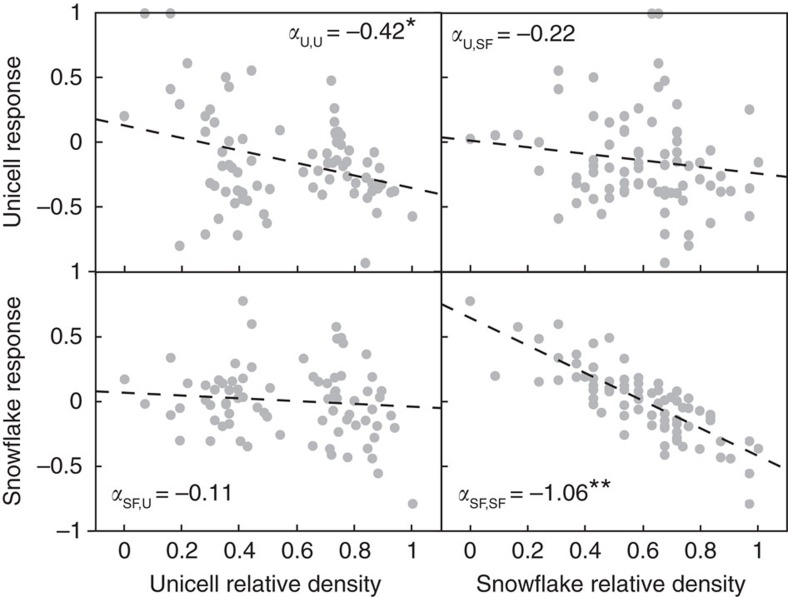
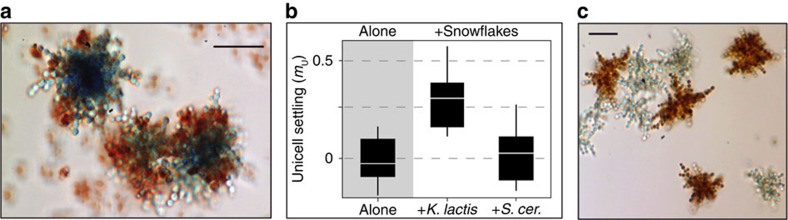
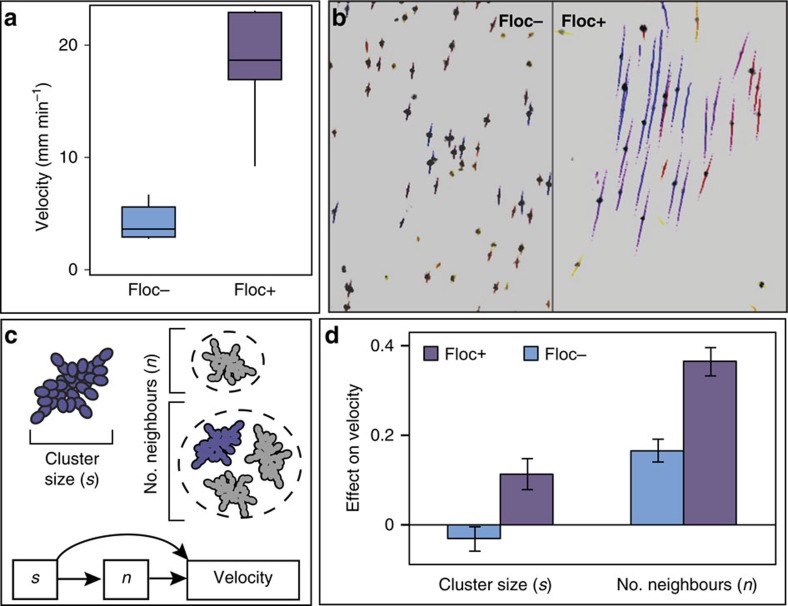
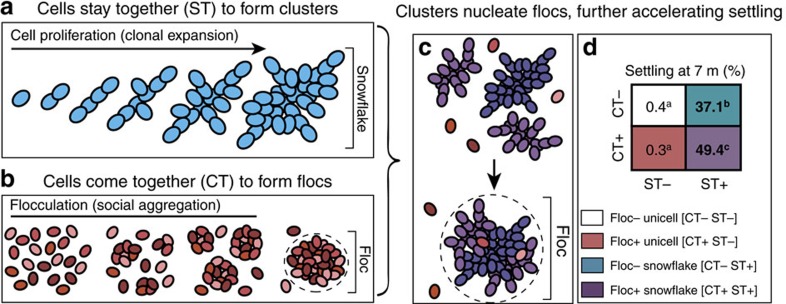
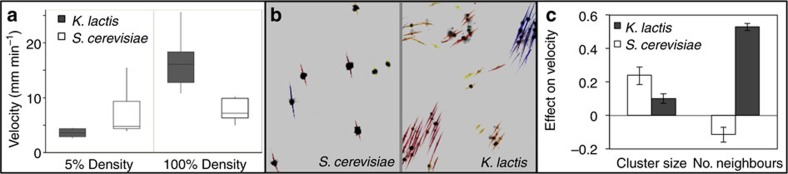
The evolution of multicellular life requires cooperation among cells, which can be undermined by intra-group selection for selfishness. Theory predicts that selection to avoid non-cooperators limits social interactions among non-relatives, yet previous evolution experiments suggest that intra-group conflict is an outcome, rather than a driver, of incipient multicellular life cycles. Here we report the evolution of multicellularity via two distinct mechanisms of group formation in the unicellular budding yeast Kluyveromyces lactis. Cells remain permanently attached following mitosis, giving rise to clonal clusters (staying together); clusters then reversibly assemble into social groups (coming together). Coming together amplifies the benefits of multicellularity and allows social clusters to collectively outperform solitary clusters. However, cooperation among non-relatives also permits fast-growing unicellular lineages to 'free-ride' during selection for increased size. Cooperation and competition for the benefits of multicellularity promote the stable coexistence of unicellular and multicellular genotypes, underscoring the importance of social and ecological context during the transition to multicellularity.
Conflict of interest statement
The authors declare no competing financial interests.
Figures
References
-
- Maynard Smith J. & Szathmaáry E. The Major Transitions in Evolution Oxford Univ. Press (1995).
-
- Merlo L. M. F., Pepper J. W., Reid B. J. & Maley C. C. Cancer as an evolutionary and ecological process. Nat. Rev. Cancer 6, 924–935 (2006). - PubMed
-
- Leigh E. G. Genes bees and ecosystems: the evolution of a common interest among individuals. Trends Ecol. Evol. 6, 257–262 (1991). - PubMed
-
- Frank S. A. Natural selection. VII. History and interpretation of kin selection theory. J. Evol. Biol. 26, 1151–1184 (2013). - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
