

NK cell heparanase controls tumor invasion and immune surveillance
- PMID: 28581441
- PMCID: PMC5490772
- DOI: 10.1172/JCI92958
NK cell heparanase controls tumor invasion and immune surveillance
Retraction in
-
NK cell heparanase controls tumor invasion and immune surveillance.J Clin Invest. 2024 Jul 1;134(13):e183295. doi: 10.1172/JCI183295. J Clin Invest. 2024. PMID: 38949030 Free PMC article. No abstract available.
Abstract
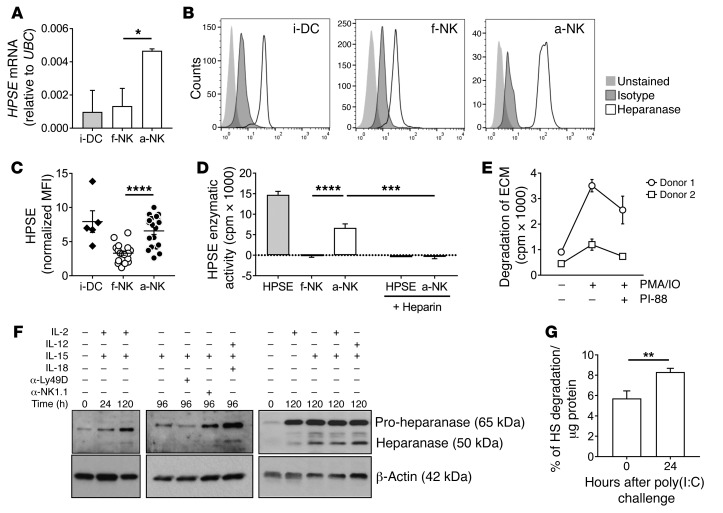
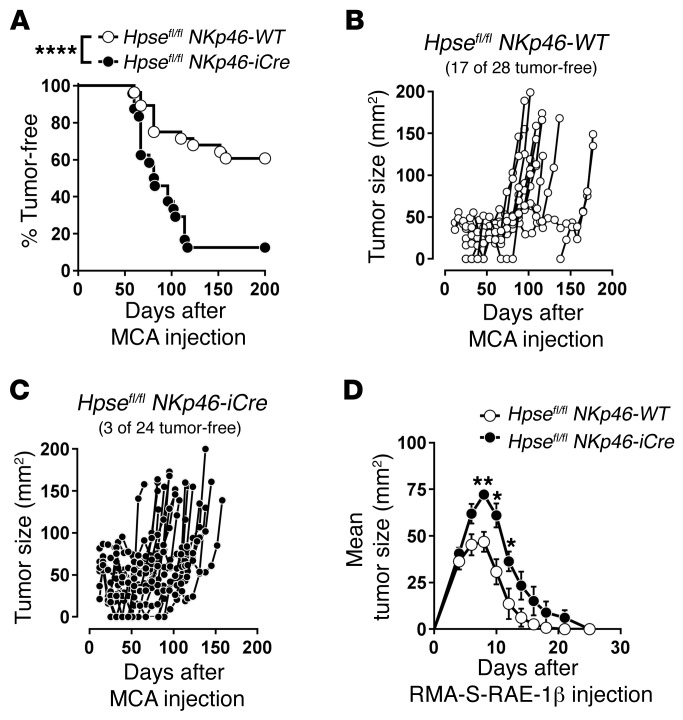
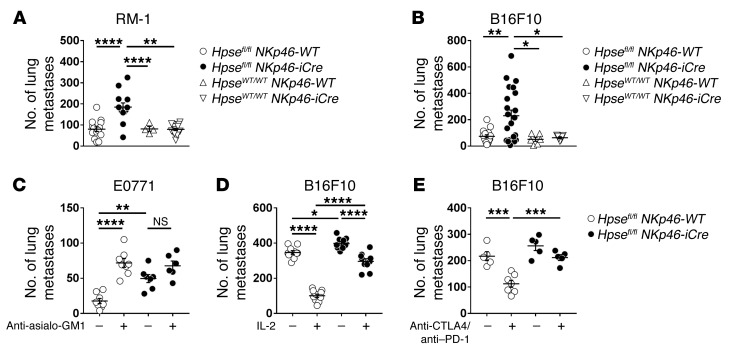
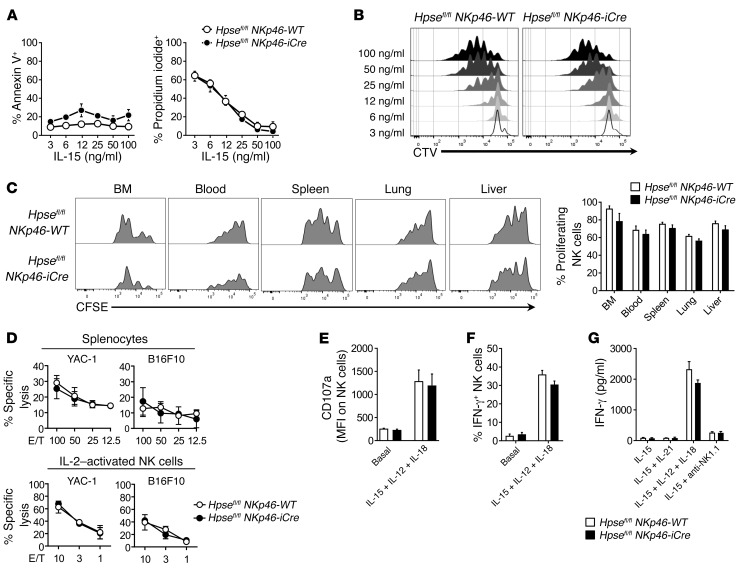
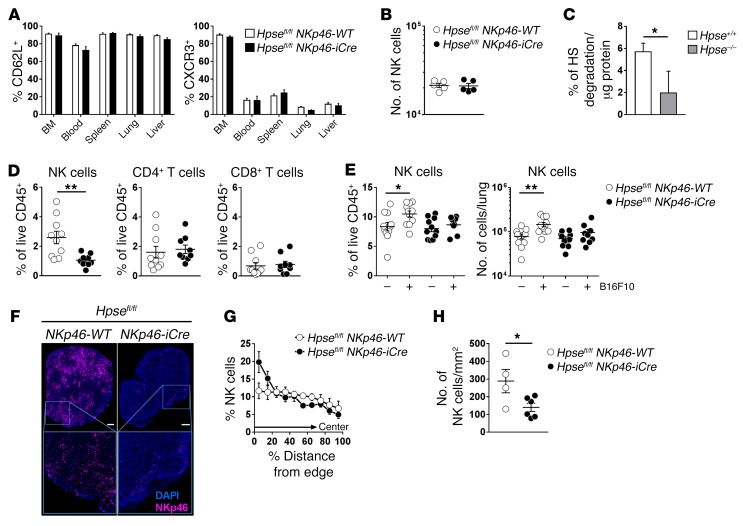
NK cells are highly efficient at preventing cancer metastasis but are infrequently found in the core of primary tumors. Here, have we demonstrated that freshly isolated mouse and human NK cells express low levels of the endo-β-D-glucuronidase heparanase that increase upon NK cell activation. Heparanase deficiency did not affect development, differentiation, or tissue localization of NK cells under steady-state conditions. However, mice lacking heparanase specifically in NK cells (Hpsefl/fl NKp46-iCre mice) were highly tumor prone when challenged with the carcinogen methylcholanthrene (MCA). Hpsefl/fl NKp46-iCre mice were also more susceptible to tumor growth than were their littermate controls when challenged with the established mouse lymphoma cell line RMA-S-RAE-1β, which overexpresses the NK cell group 2D (NKG2D) ligand RAE-1β, or when inoculated with metastatic melanoma, prostate carcinoma, or mammary carcinoma cell lines. NK cell invasion of primary tumors and recruitment to the site of metastasis were strictly dependent on the presence of heparanase. Cytokine and immune checkpoint blockade immunotherapy for metastases was compromised when NK cells lacked heparanase. Our data suggest that heparanase plays a critical role in NK cell invasion into tumors and thereby tumor progression and metastases. This should be considered when systemically treating cancer patients with heparanase inhibitors, since the potential adverse effect on NK cell infiltration might limit the antitumor activity of the inhibitors.
Conflict of interest statement
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
