

Regulation of cargo transfer between ESCRT-0 and ESCRT-I complexes by flotillin-1 during endosomal sorting of ubiquitinated cargo
- PMID: 28581508
- PMCID: PMC5519196
- DOI: 10.1038/oncsis.2017.47
Regulation of cargo transfer between ESCRT-0 and ESCRT-I complexes by flotillin-1 during endosomal sorting of ubiquitinated cargo
Abstract
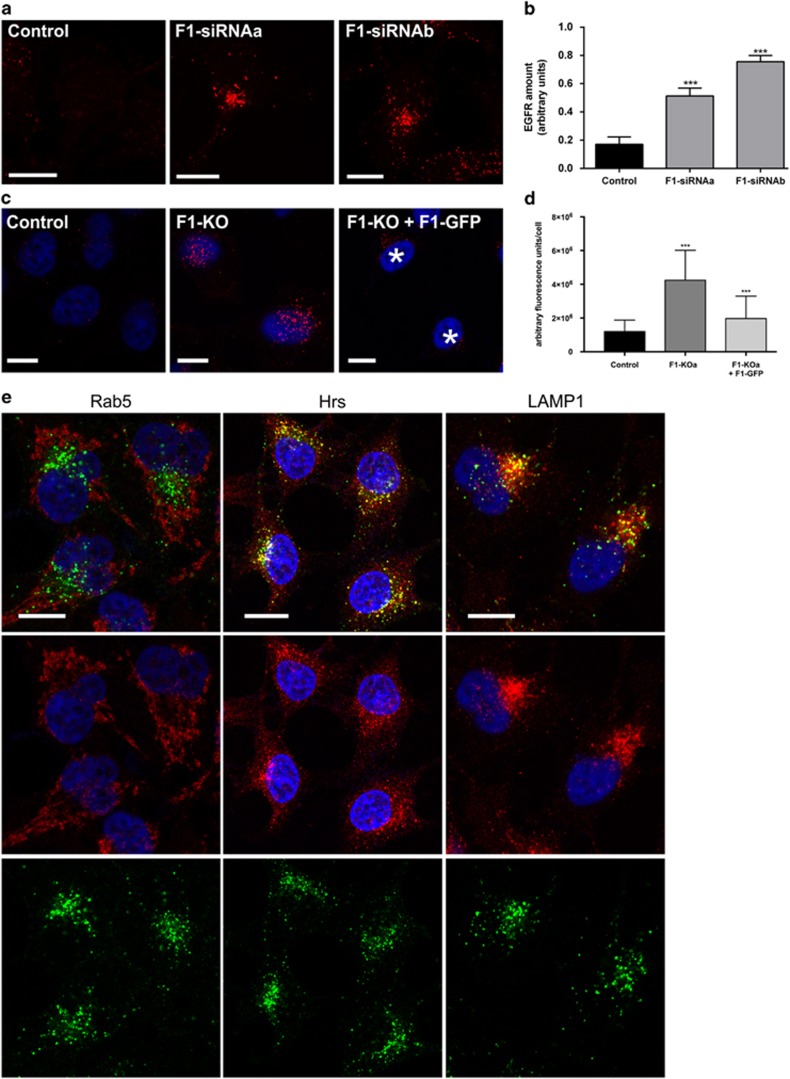
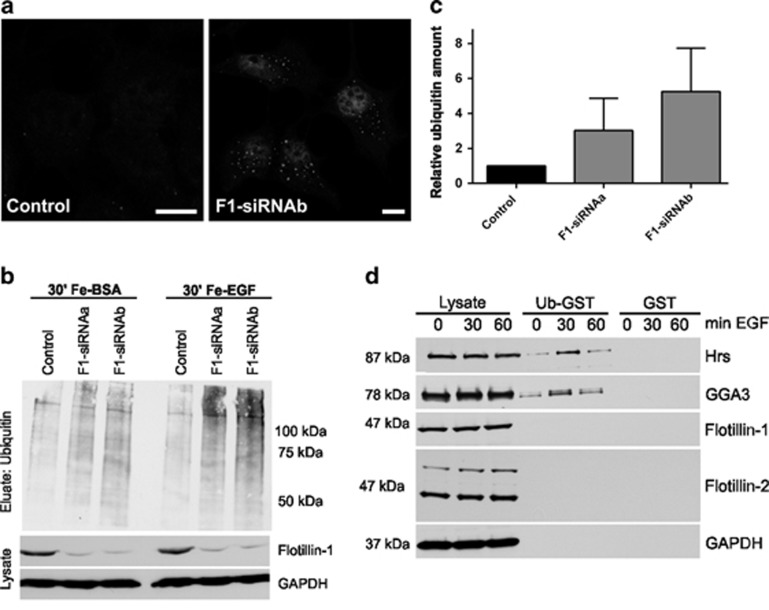
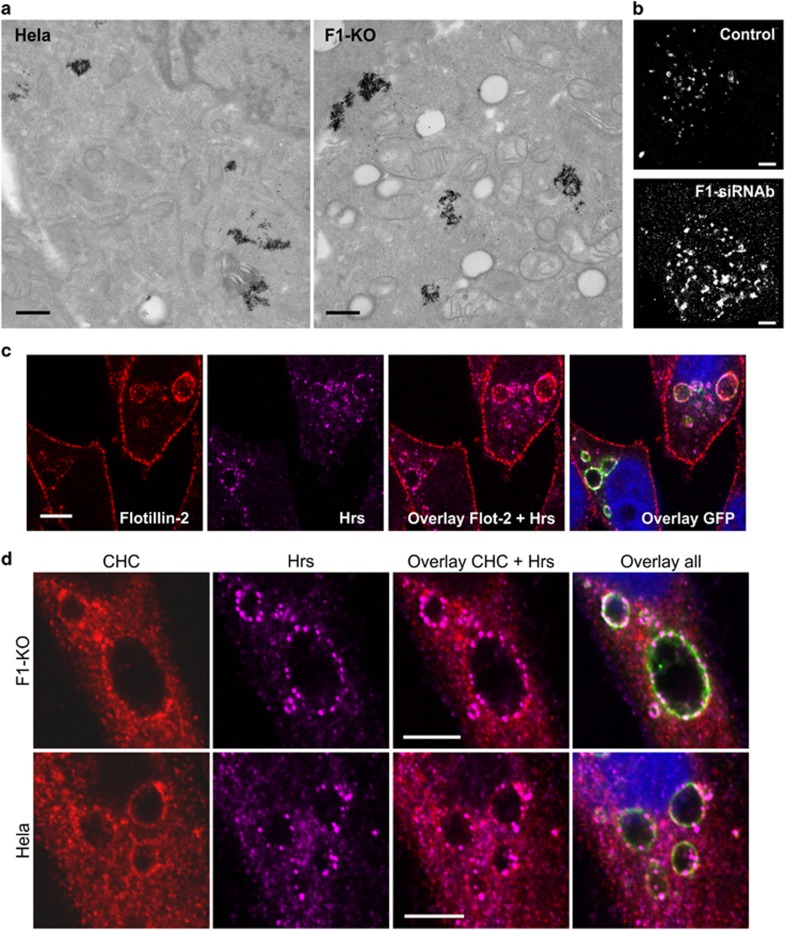
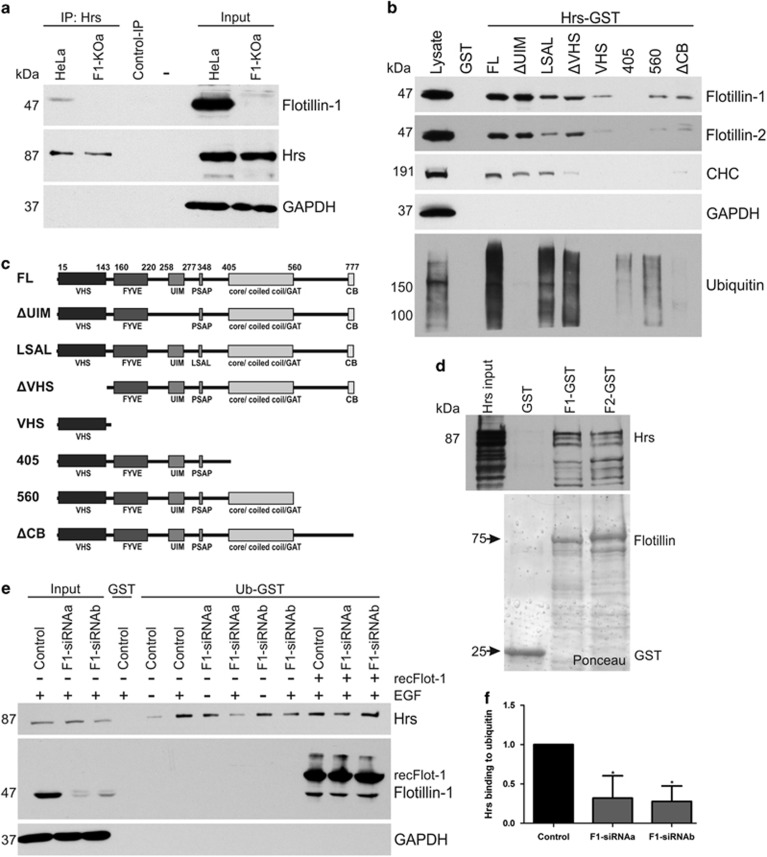
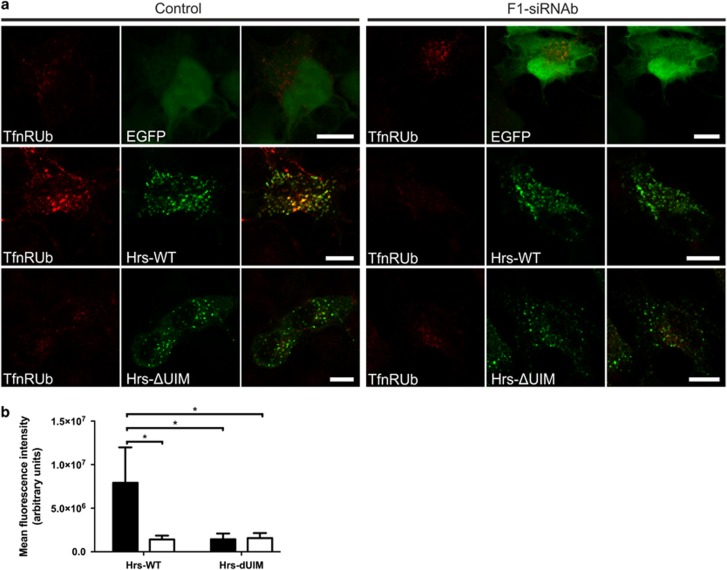
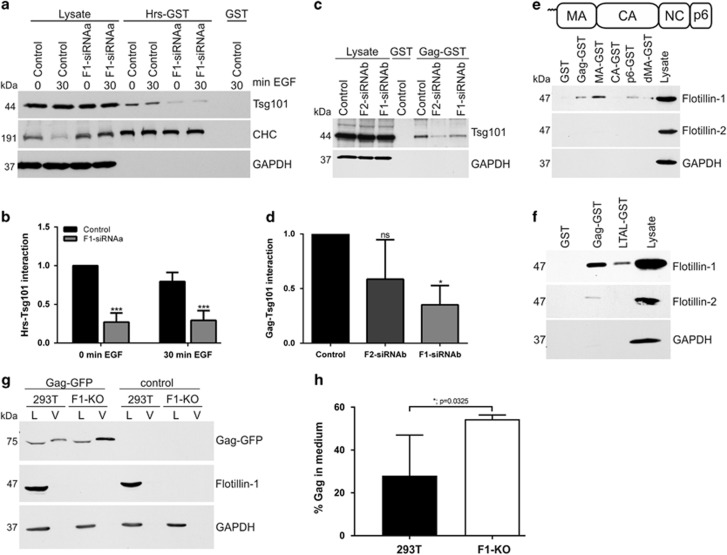
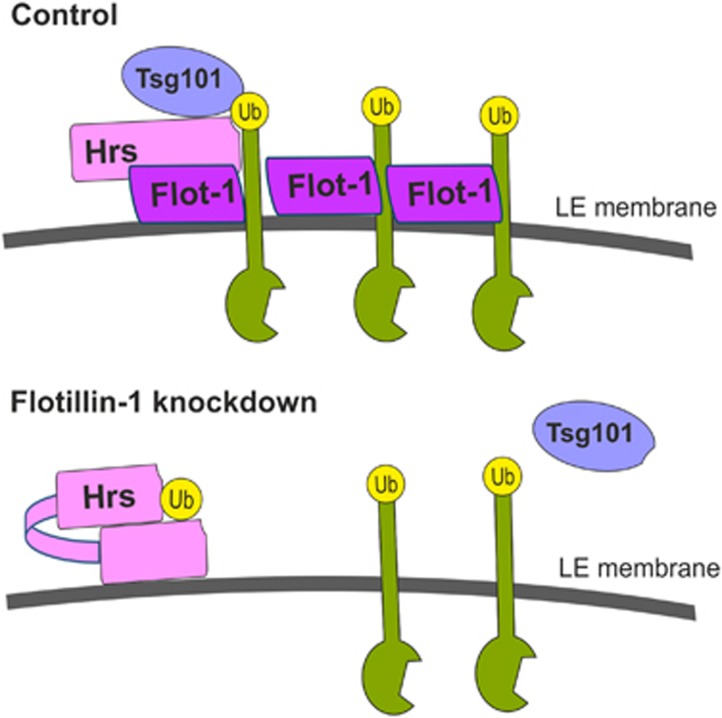
Ubiquitin-dependent sorting of membrane proteins in endosomes directs them to lysosomal degradation. In the case of receptors such as the epidermal growth factor receptor (EGFR), lysosomal degradation is important for the regulation of downstream signalling. Ubiquitinated proteins are recognised in endosomes by the endosomal sorting complexes required for transport (ESCRT) complexes, which sequentially interact with the ubiquitinated cargo. Although the role of each ESCRT complex in sorting is well established, it is not clear how the cargo is passed on from one ESCRT to the next. We here show that flotillin-1 is required for EGFR degradation, and that it interacts with the subunits of ESCRT-0 and -I complexes (hepatocyte growth factor-regulated tyrosine kinase substrate (Hrs) and Tsg101). Flotillin-1 is required for cargo recognition and sorting by ESCRT-0/Hrs and for its interaction with Tsg101. In addition, flotillin-1 is also required for the sorting of human immunodeficiency virus 1 Gag polyprotein, which mimics ESCRT-0 complex during viral assembly. We propose that flotillin-1 functions in cargo transfer between ESCRT-0 and -I complexes.
Conflict of interest statement
The authors declare no conflict of interest.
Figures

References
-
- Henne WM, Buchkovich NJ, Emr SD. The ESCRT pathway. Dev Cell 2011; 21: 77–91. - PubMed
-
- Katzmann DJ, Babst M, Emr SD. Ubiquitin-dependent sorting into the multivesicular body pathway requires the function of a conserved endosomal protein sorting complex ESCRT-I. Cell 2001; 106: 145–155. - PubMed
-
- Raiborg C, Bremnes B, Mehlum A, Gillooly DJ, D'Arrigo A, Stang E et al. FYVE and coiled-coil domains determine the specific localisation of Hrs to early endosomes. J Cell Sci 2001; 114(Part 12): 2255–2263. - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
