

Proteasome Structure and Assembly
- PMID: 28583440
- PMCID: PMC5675778
- DOI: 10.1016/j.jmb.2017.05.027
Proteasome Structure and Assembly
Abstract
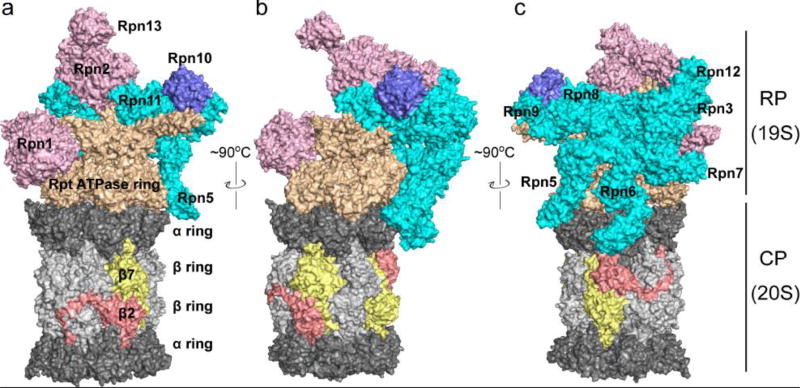
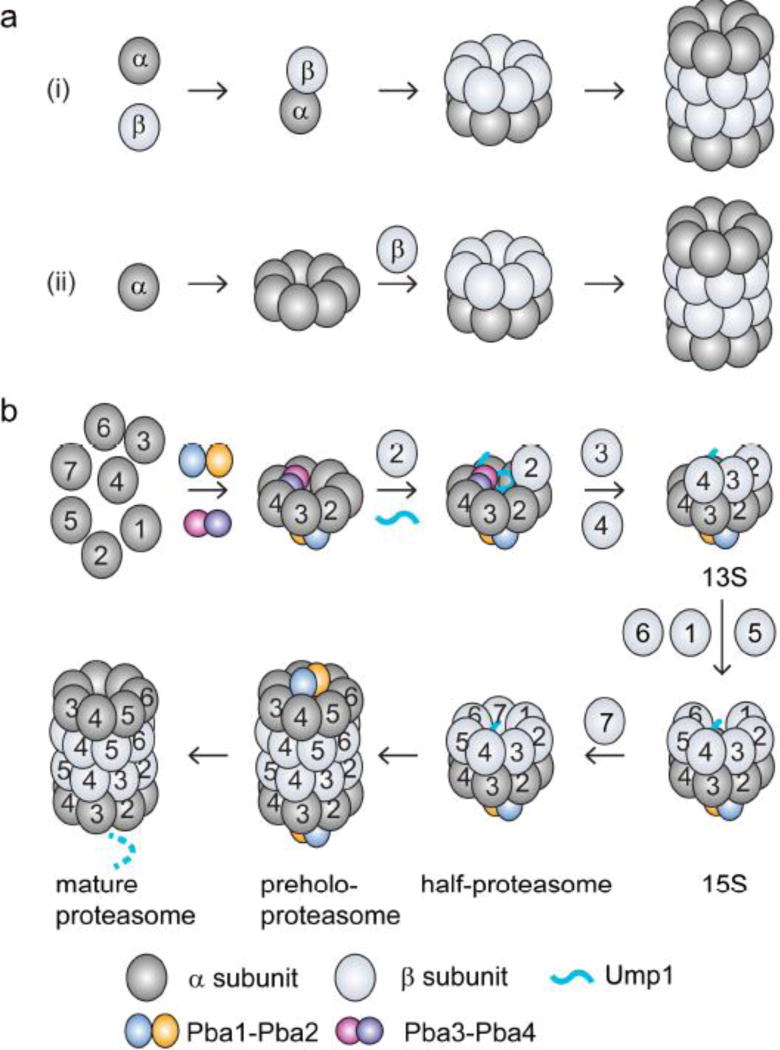
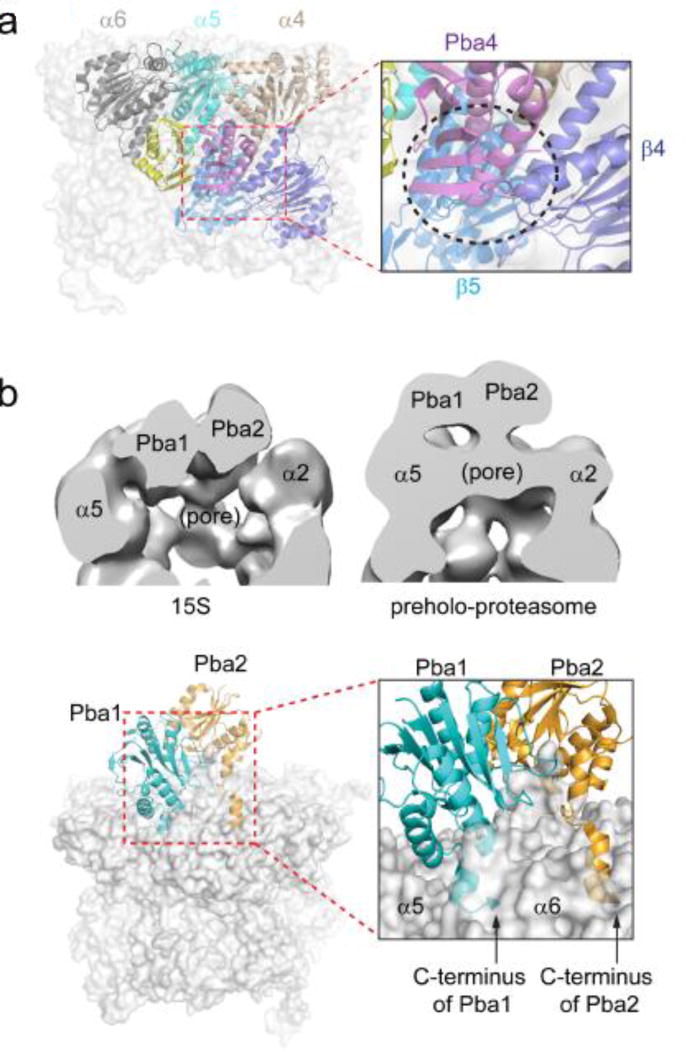
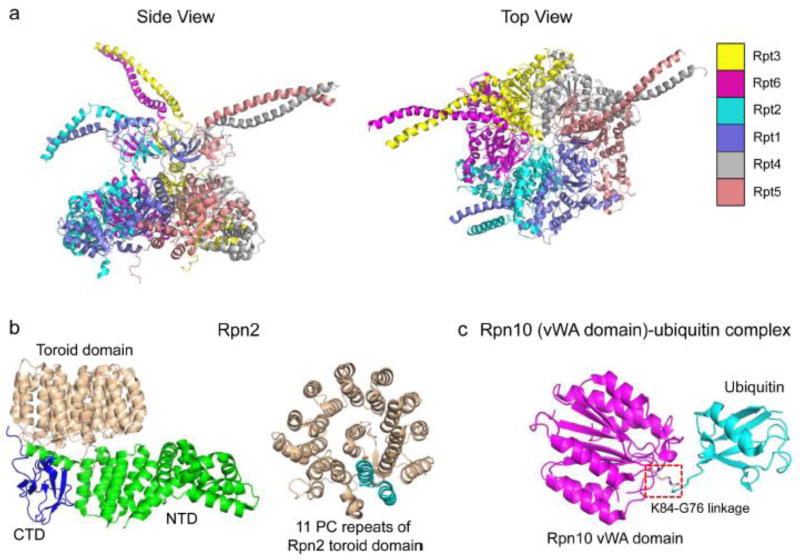
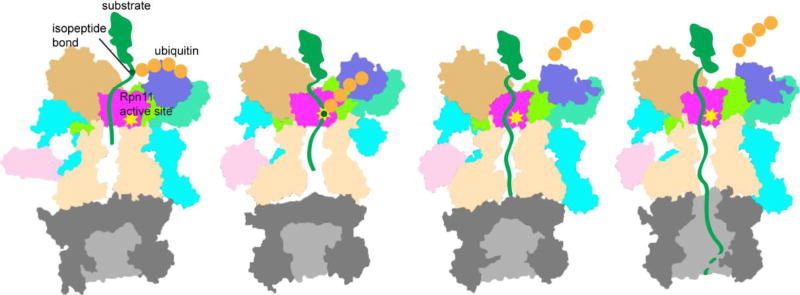
The eukaryotic 26S proteasome is a large multisubunit complex that degrades the majority of proteins in the cell under normal conditions. The 26S proteasome can be divided into two subcomplexes: the 19S regulatory particle and the 20S core particle. Most substrates are first covalently modified by ubiquitin, which then directs them to the proteasome. The function of the regulatory particle is to recognize, unfold, deubiquitylate, and translocate substrates into the core particle, which contains the proteolytic sites of the proteasome. Given the abundance and subunit complexity of the proteasome, the assembly of this ~2.5MDa complex must be carefully orchestrated to ensure its correct formation. In recent years, significant progress has been made in the understanding of proteasome assembly, structure, and function. Technical advances in cryo-electron microscopy have resulted in a series of atomic cryo-electron microscopy structures of both human and yeast 26S proteasomes. These structures have illuminated new intricacies and dynamics of the proteasome. In this review, we focus on the mechanisms of proteasome assembly, particularly in light of recent structural information.
Keywords: proteasome; proteasome assembly; protein degradation; ubiquitin proteasome system.
Copyright © 2017 Elsevier Ltd. All rights reserved.
Figures
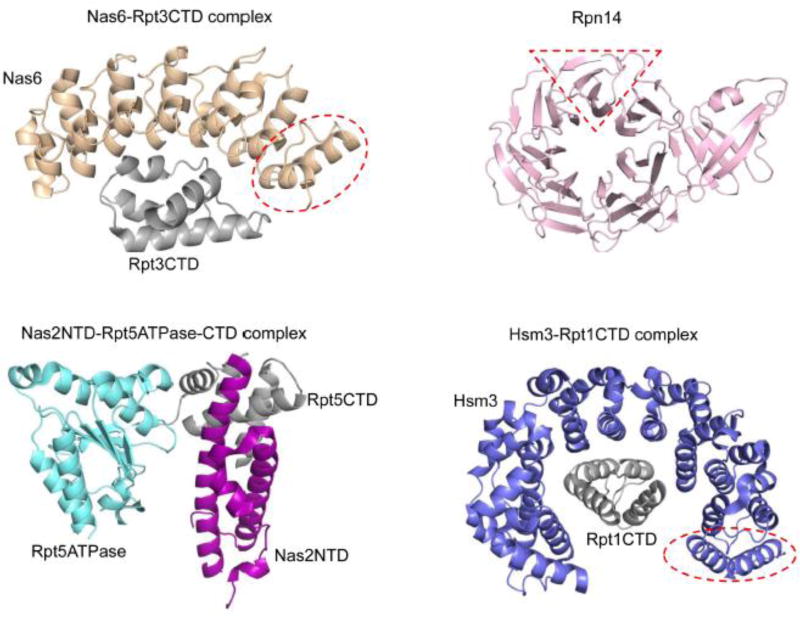
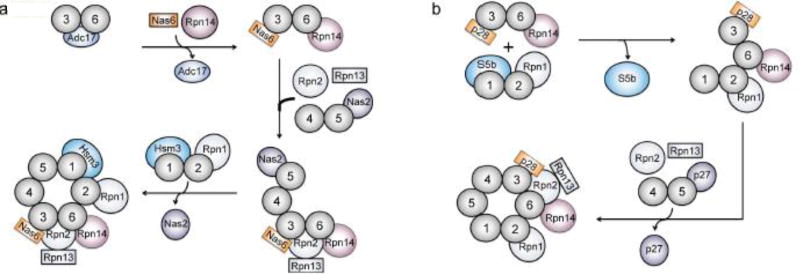
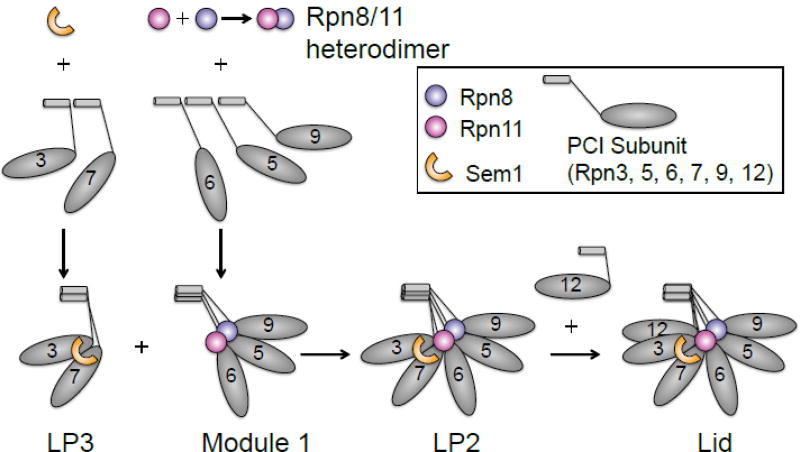
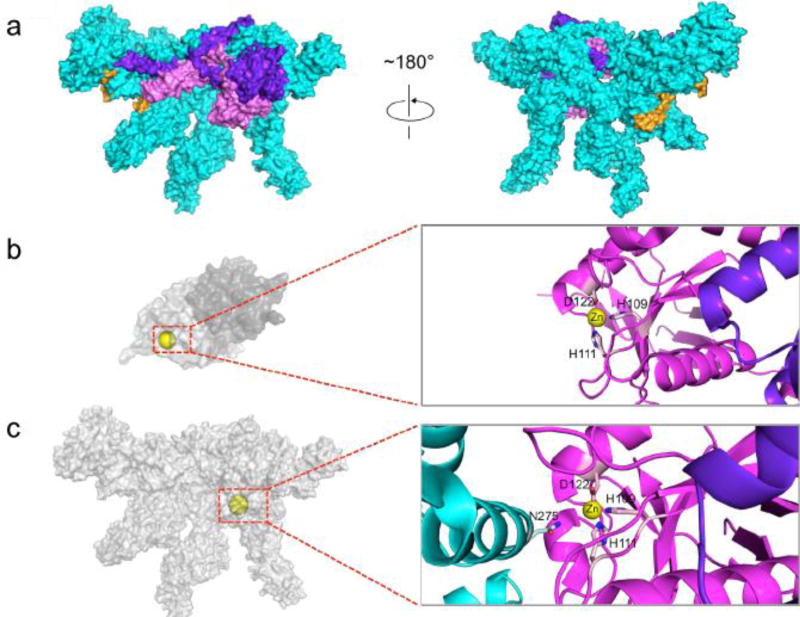
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
