

Numerosity processing in early visual cortex
- PMID: 28583882
- PMCID: PMC6697050
- DOI: 10.1016/j.neuroimage.2017.05.069
Numerosity processing in early visual cortex
Abstract
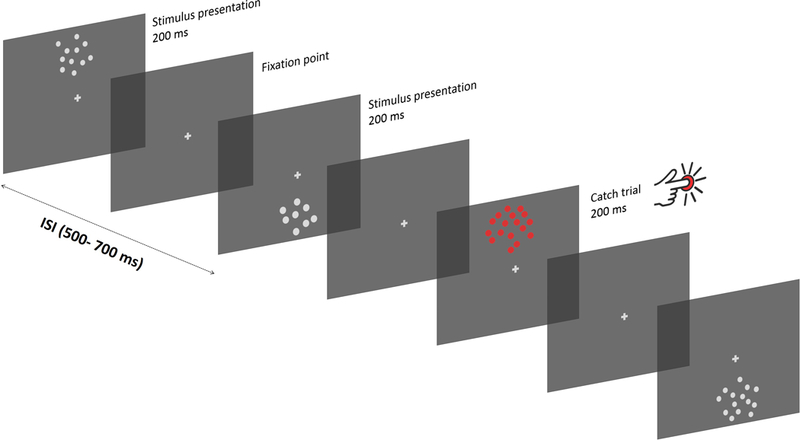
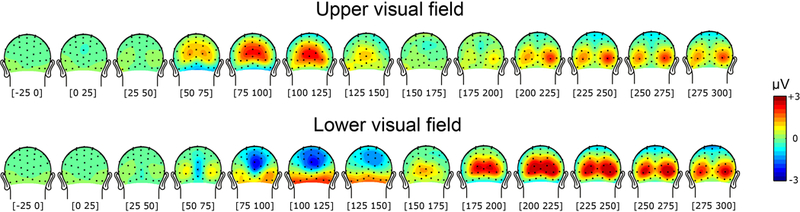
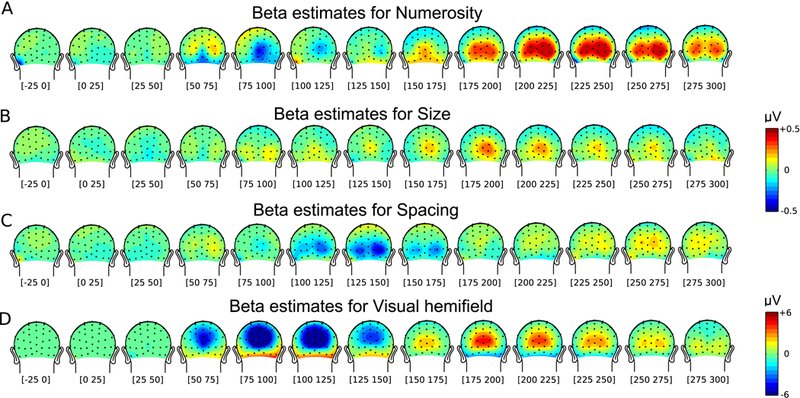
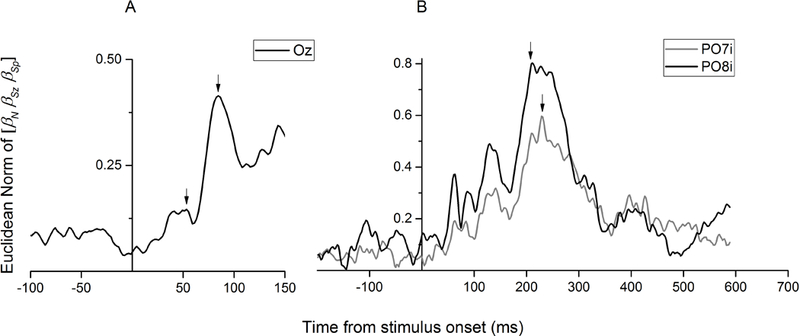
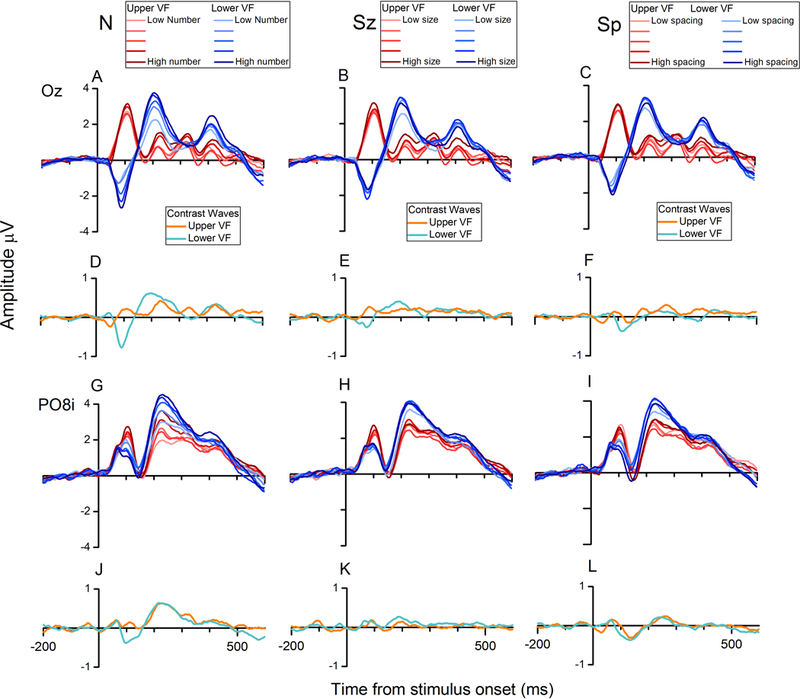
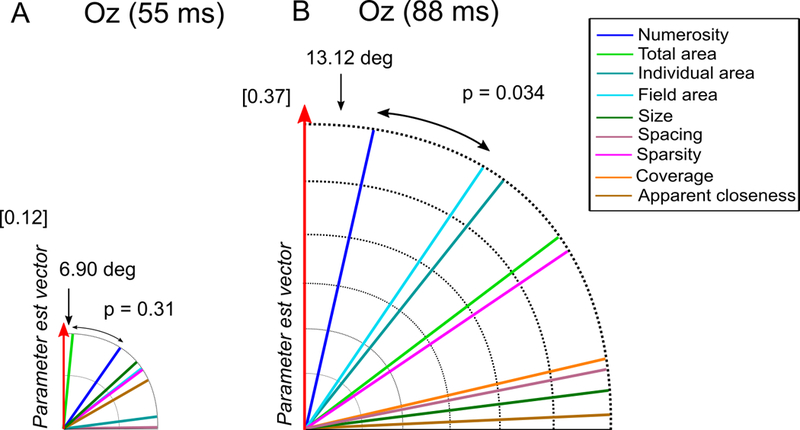
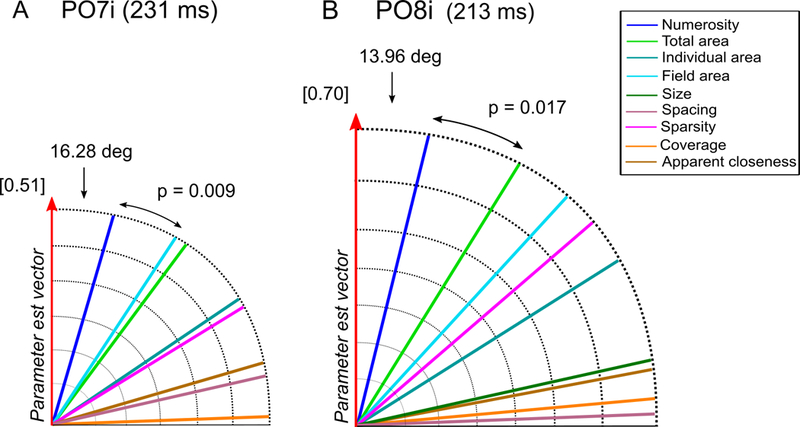
While parietal cortex is thought to be critical for representing numerical magnitudes, we recently reported an event-related potential (ERP) study demonstrating selective neural sensitivity to numerosity over midline occipital sites very early in the time course, suggesting the involvement of early visual cortex in numerosity processing. However, which specific brain area underlies such early activation is not known. Here, we tested whether numerosity-sensitive neural signatures arise specifically from the initial stages of visual cortex, aiming to localize the generator of these signals by taking advantage of the distinctive folding pattern of early occipital cortices around the calcarine sulcus, which predicts an inversion of polarity of ERPs arising from these areas when stimuli are presented in the upper versus lower visual field. Dot arrays, including 8-32dots constructed systematically across various numerical and non-numerical visual attributes, were presented randomly in either the upper or lower visual hemifields. Our results show that neural responses at about 90ms post-stimulus were robustly sensitive to numerosity. Moreover, the peculiar pattern of polarity inversion of numerosity-sensitive activity at this stage suggested its generation primarily in V2 and V3. In contrast, numerosity-sensitive ERP activity at occipito-parietal channels later in the time course (210-230ms) did not show polarity inversion, indicating a subsequent processing stage in the dorsal stream. Overall, these results demonstrate that numerosity processing begins in one of the earliest stages of the cortical visual stream.
Keywords: Calcarine sulcus; ERPs; Numerosity; Visual cortex; Visual processing.
Copyright © 2017 Elsevier Inc. All rights reserved.
Figures
Similar articles
-
Early Numerosity Encoding in Visual Cortex Is Not Sufficient for the Representation of Numerical Magnitude.J Cogn Neurosci. 2018 Dec;30(12):1788-1802. doi: 10.1162/jocn_a_01320. Epub 2018 Jul 31. J Cogn Neurosci. 2018. PMID: 30063175
-
Rapid and Direct Encoding of Numerosity in the Visual Stream.Cereb Cortex. 2016 Feb;26(2):748-763. doi: 10.1093/cercor/bhv017. Epub 2015 Feb 24. Cereb Cortex. 2016. PMID: 25715283 Free PMC article.
-
Behavior-related potentials from single-trial interindividual correlation between event related potentials and behavioral performance reveals right lateralized processing of numerosity.Brain Cogn. 2024 Oct;180:106185. doi: 10.1016/j.bandc.2024.106185. Epub 2024 Jun 15. Brain Cogn. 2024. PMID: 38878607
-
Distributed and interactive brain mechanisms during emotion face perception: evidence from functional neuroimaging.Neuropsychologia. 2007 Jan 7;45(1):174-94. doi: 10.1016/j.neuropsychologia.2006.06.003. Epub 2006 Jul 18. Neuropsychologia. 2007. PMID: 16854439 Review.
-
Revealing the body in the brain: An ERP method to examine sensorimotor activity during visual perception of body-related information.Cortex. 2020 Apr;125:332-344. doi: 10.1016/j.cortex.2020.01.017. Epub 2020 Feb 6. Cortex. 2020. PMID: 32120169 Review.
Cited by
-
Fast saccadic eye-movements in humans suggest that numerosity perception is automatic and direct.Proc Biol Sci. 2020 Sep 30;287(1935):20201884. doi: 10.1098/rspb.2020.1884. Epub 2020 Sep 23. Proc Biol Sci. 2020. PMID: 32962551 Free PMC article.
-
The neural correlates of individual differences in numerosity perception: A voxel-based morphometry study.iScience. 2023 Jul 14;26(8):107392. doi: 10.1016/j.isci.2023.107392. eCollection 2023 Aug 18. iScience. 2023. PMID: 37554464 Free PMC article.
-
Perceptual History Biases Are Predicted by Early Visual-Evoked Activity.J Neurosci. 2023 May 24;43(21):3860-3875. doi: 10.1523/JNEUROSCI.1451-22.2023. Epub 2023 Apr 21. J Neurosci. 2023. PMID: 37085319 Free PMC article.
-
Serial dependence in time and numerosity perception is dimension-specific.J Vis. 2021 May 3;21(5):6. doi: 10.1167/jov.21.5.6. J Vis. 2021. PMID: 33956059 Free PMC article.
-
Shape facilitates number: brain potentials and microstates reveal the interplay between shape and numerosity in human vision.Sci Rep. 2020 Jul 24;10(1):12413. doi: 10.1038/s41598-020-68788-4. Sci Rep. 2020. PMID: 32709892 Free PMC article.
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
