

Allosteric modulation of human dopamine transporter activity under conditions promoting its dimerization
- PMID: 28584050
- PMCID: PMC5535022
- DOI: 10.1074/jbc.M116.763565
Allosteric modulation of human dopamine transporter activity under conditions promoting its dimerization
Abstract
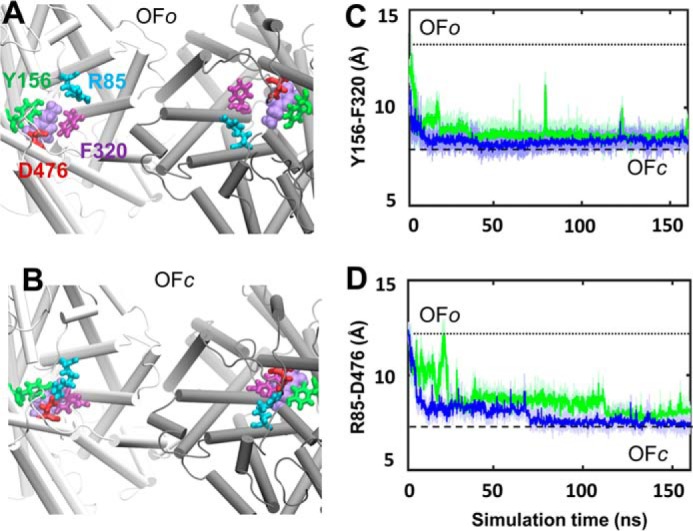
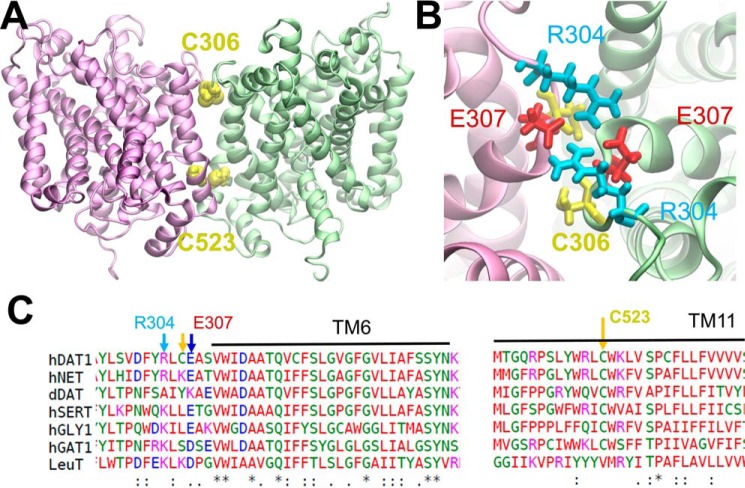
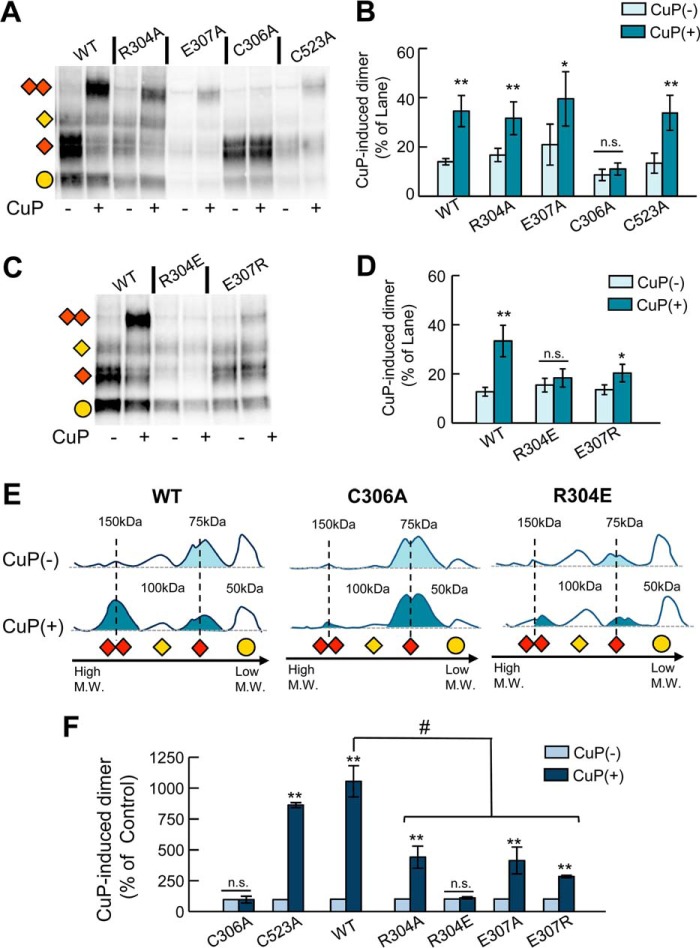
The human dopamine (DA) transporter (hDAT) is a key regulator of neurotransmission and a target for antidepressants and addictive drugs. Despite the recent resolution of dDAT structures from Drosophila melanogaster, complete understanding of its mechanism of function and even information on its biological assembly is lacking. The resolved dDAT structures are monomeric, but growing evidence suggests that hDAT might function as a multimer, and its oligomerization may be relevant to addictive drug effects. Here, using structure-based computations, we examined the possible mechanisms of hDAT dimerization and its dynamics in a lipid bilayer. Using a combination of site-directed mutagenesis, DA-uptake, and cross-linking experiments that exploited the capacity of Cys-306 to form intermonomeric disulfide bridges in the presence of an oxidizing agent, we tested the effects of mutations at transmembrane segment (TM) 6 and 12 helices in HEK293 cells. The most probable structural model for hDAT dimer suggested by computations and experiments differed from the dimeric structure resolved for the bacterial homolog, LeuT, presumably because of a kink at TM12 preventing favorable monomer packing. Instead, TM2, TM6, and TM11 line the dimer interface. Molecular dynamics simulations of the dimeric hDAT indicated that the two subunits tend to undergo cooperative structural changes, both on local (extracellular gate opening/closure) and global (transition between outward-facing and inward-facing states) scales. These observations suggest that hDAT transport properties may be allosterically modulated under conditions promoting dimerization. Our study provides critical insights into approaches for examining the oligomerization of neurotransmitter transporters and sheds light on their drug modulation.
Keywords: docking; dopamine transporter; molecular dynamics; mutagenesis; protein assembly.
© 2017 by The American Society for Biochemistry and Molecular Biology, Inc.
Conflict of interest statement
The authors declare that they have no conflicts of interest with the contents of this article
Figures
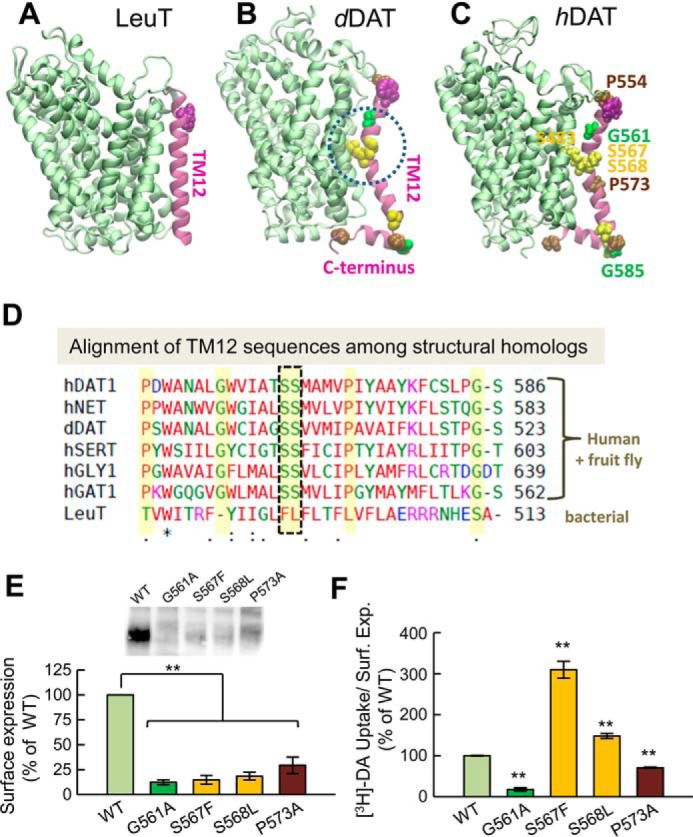
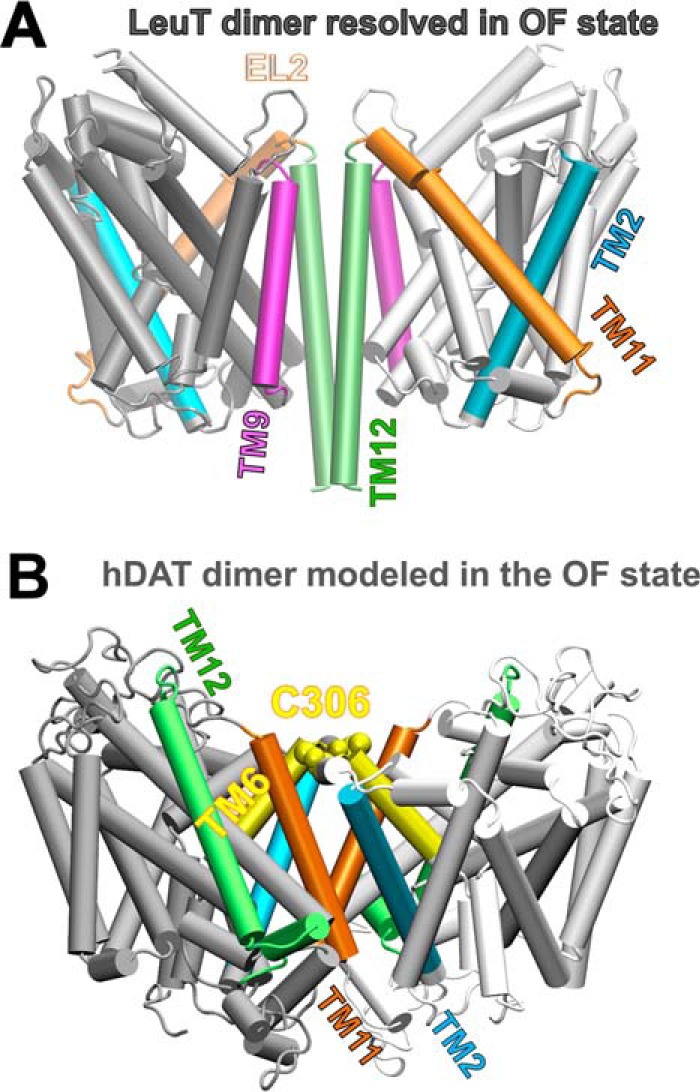
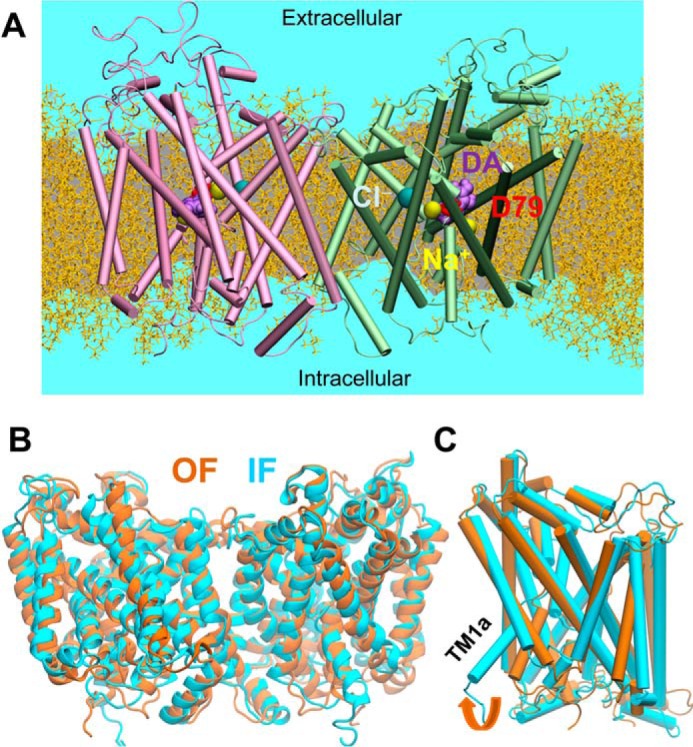
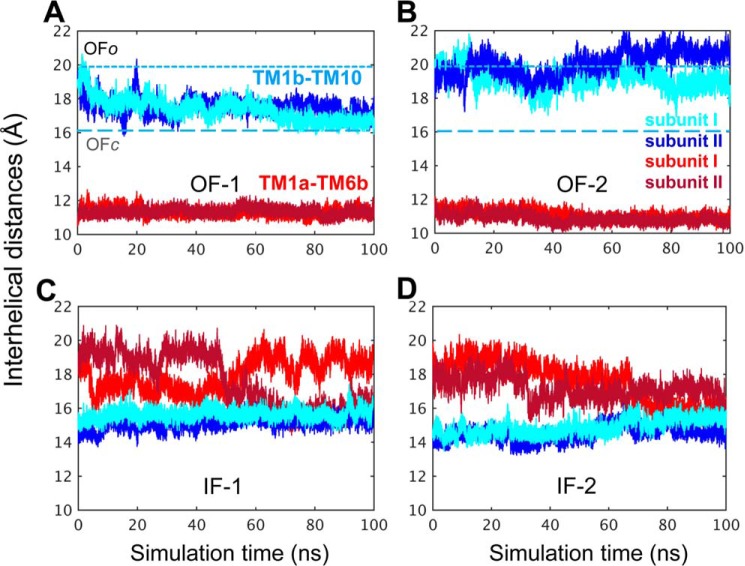
References
-
- Amara S. G., and Sonders M. S. (1998) Neurotransmitter transporters as molecular targets for addictive drugs. Drug Alcohol Depend. 51, 87–96 - PubMed
-
- Reith M. (2002) Neurotransmitter transporters: Structure, Function, and Regulations. Humana, Totowa, NJ
-
- Jardetzky O. (1966) Simple allosteric model for membrane pumps. Nature 211, 969–970 - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
