

Genomic Insights into the Evolution of the Nicotine Biosynthesis Pathway in Tobacco
- PMID: 28584068
- PMCID: PMC5462024
- DOI: 10.1104/pp.17.00070
Genomic Insights into the Evolution of the Nicotine Biosynthesis Pathway in Tobacco
Abstract
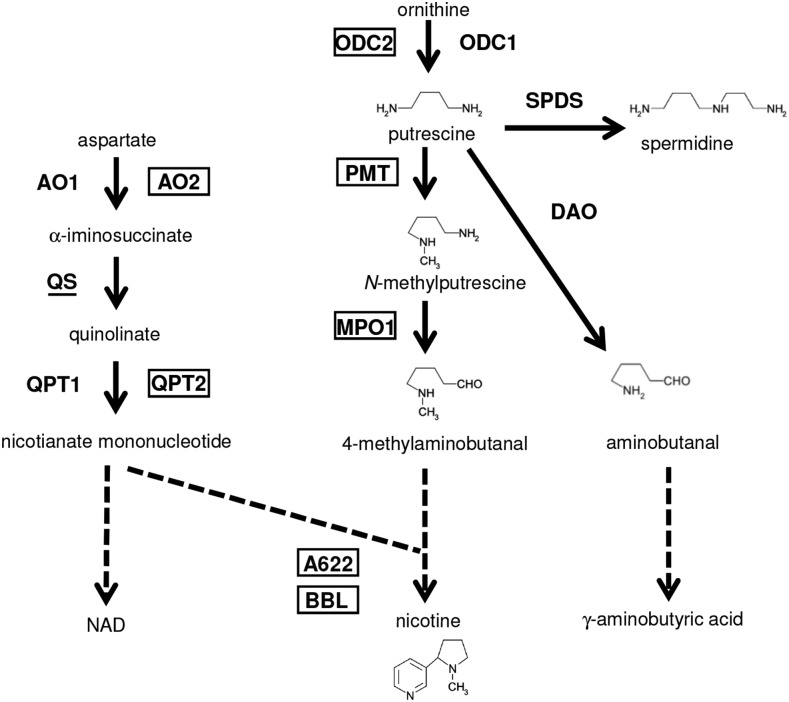
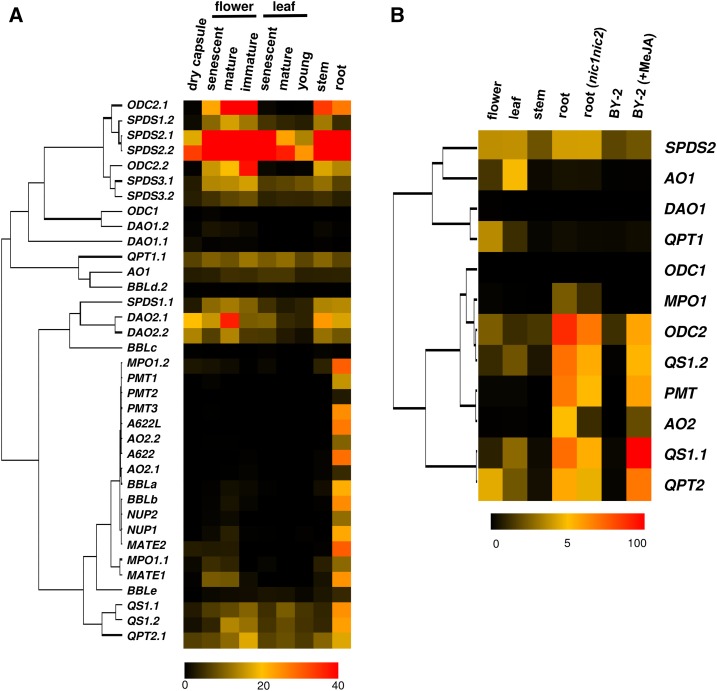
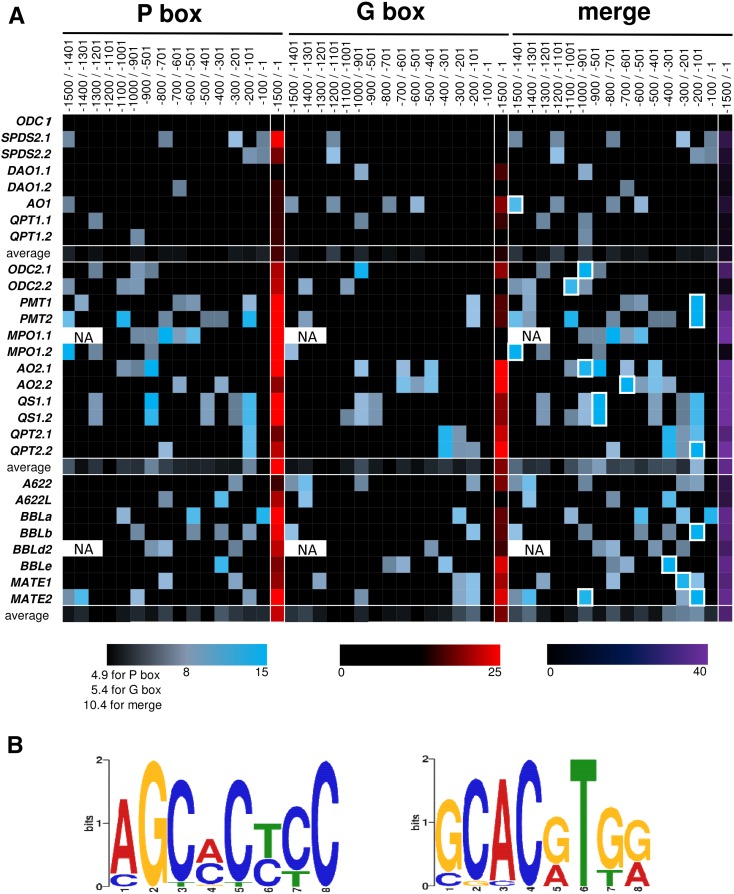
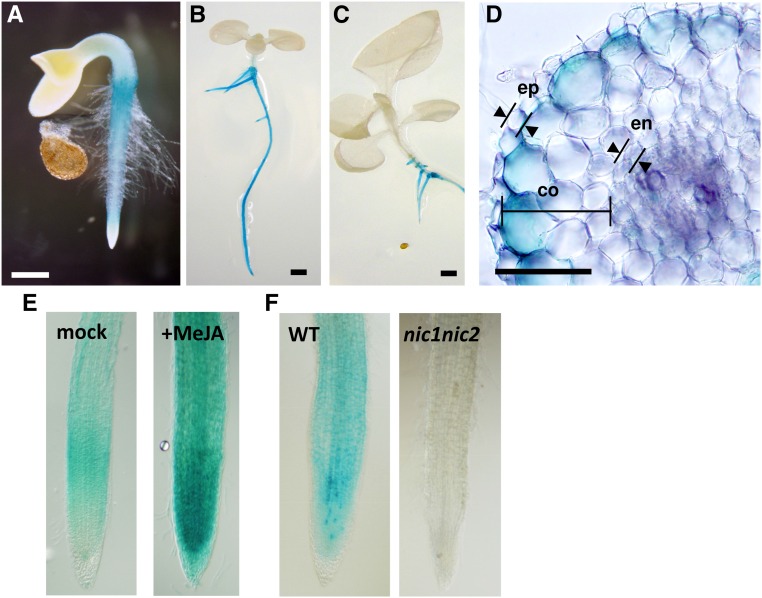
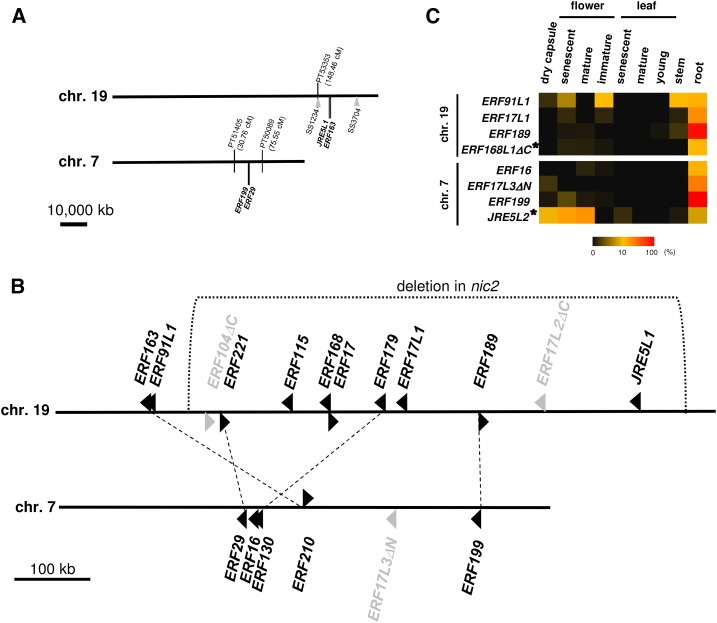
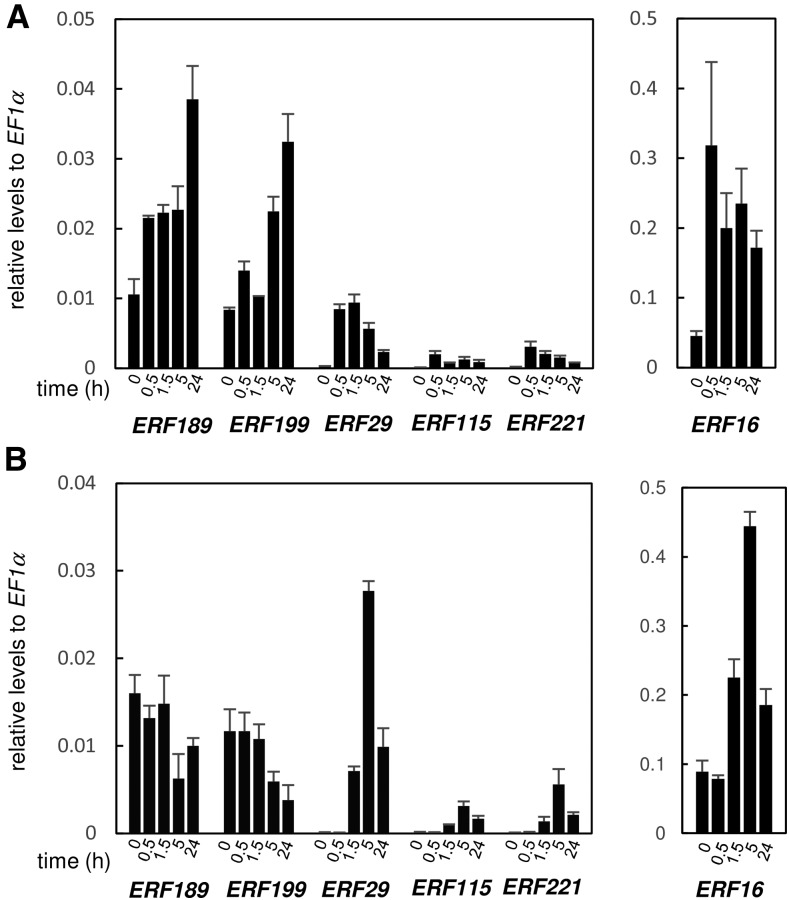
In tobacco (Nicotiana tabacum), nicotine is the predominant alkaloid. It is produced in the roots and accumulated mainly in the leaves. Jasmonates play a central signaling role in damage-induced nicotine formation. The genome sequence of tobacco provides us an almost complete inventory of structural and regulatory genes involved in nicotine pathway. Phylogenetic and expression analyses revealed a series of structural genes of the nicotine pathway, forming a regulon, under the control of jasmonate-responsive ETHYLENE RESPONSE FACTOR (ERF) transcription factors. The duplication of NAD and polyamine metabolic pathways and the subsequent recruitment of duplicated primary metabolic genes into the nicotine biosynthesis regulon were suggested to be the drivers for pyridine and pyrrolidine ring formation steps early in the pathway. Transcriptional regulation by ERF and cooperatively acting MYC2 transcription factors are corroborated by the frequent occurrence of cognate cis-regulatory elements of the factors in the promoter regions of the downstream structural genes. The allotetraploid tobacco has homologous clusters of ERF genes on different chromosomes, which are possibly derived from two ancestral diploids and include either nicotine-controlling ERF189 or ERF199 A large chromosomal deletion was found within one allele of the nicotine-controlling NICOTINE2 locus, which is part of one of the ERF gene clusters, and which has been used to breed tobacco cultivars with a low-nicotine content.
© 2017 The author(s). All Rights Reserved.
Figures
References
-
- Arimura G, Maffei M, editors (2017) Plant Specialized Metabolism: Genomics, Biochemistry, and Biological Functions. CRC Press, New York
-
- Baldwin IT. (1989) Mechanism of damage-induced alkaloid production in wild tobacco. J Chem Ecol 15: 1661–1680 - PubMed
-
- Baldwin IT, Schmelz EA, Ohnmeiss TE (1994) Wound-induced changes in root and shoot jasmonic acid pools correlate with induced nicotine synthesis in Nicotiana sylvestris spegazzini and comes. J Chem Ecol 20: 2139–2157 - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
