

Deubiquitinating enzyme VCIP135 dictates the duration of botulinum neurotoxin type A intoxication
- PMID: 28584101
- PMCID: PMC5495235
- DOI: 10.1073/pnas.1621076114
Deubiquitinating enzyme VCIP135 dictates the duration of botulinum neurotoxin type A intoxication
Abstract
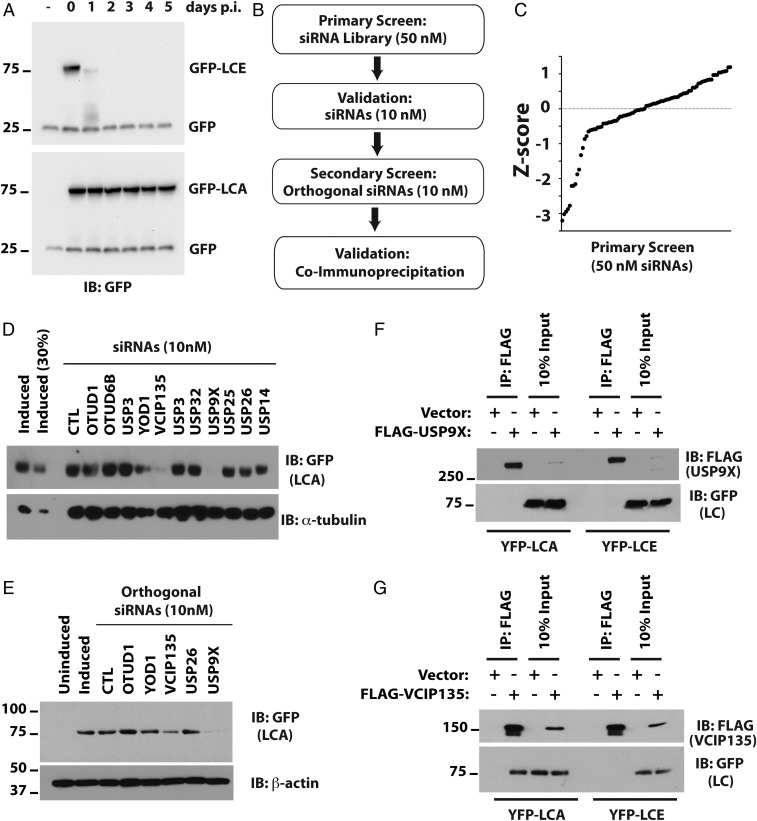
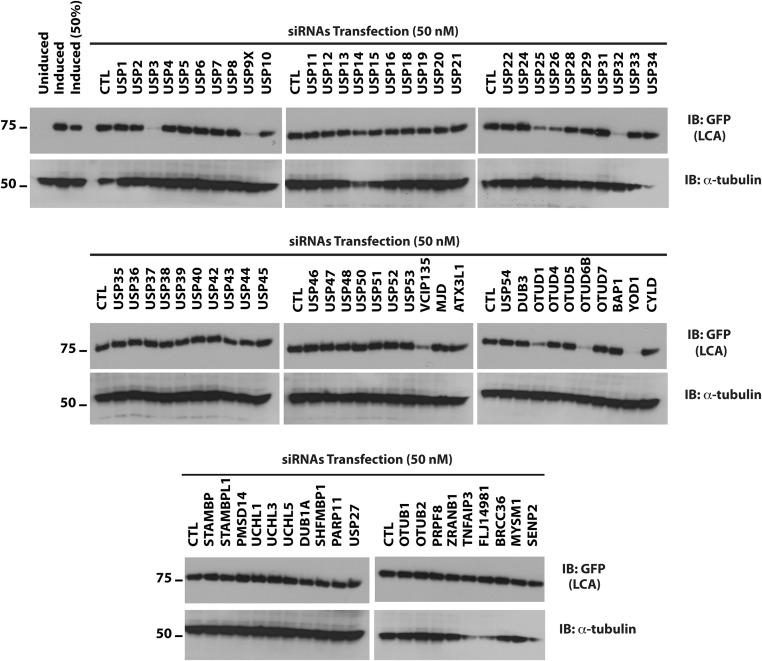
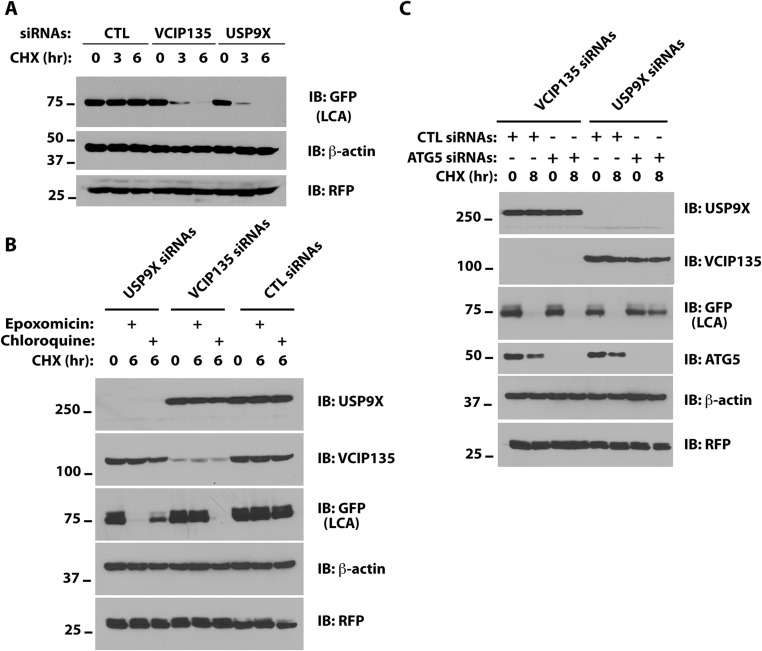
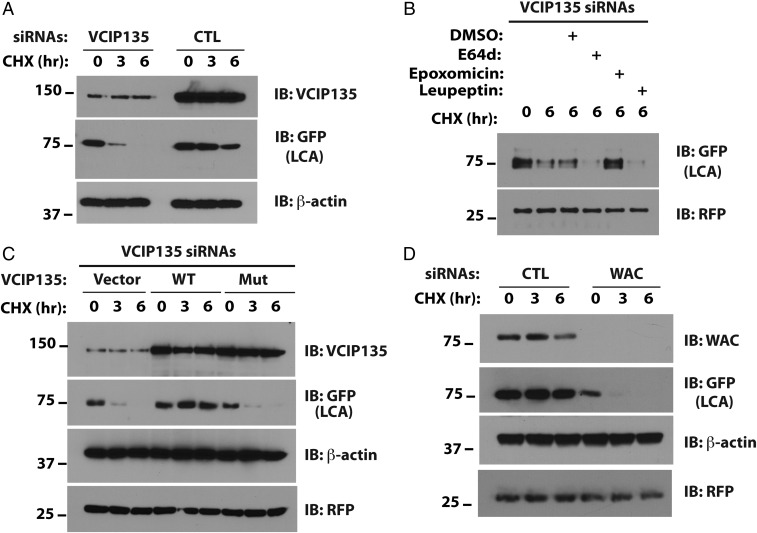
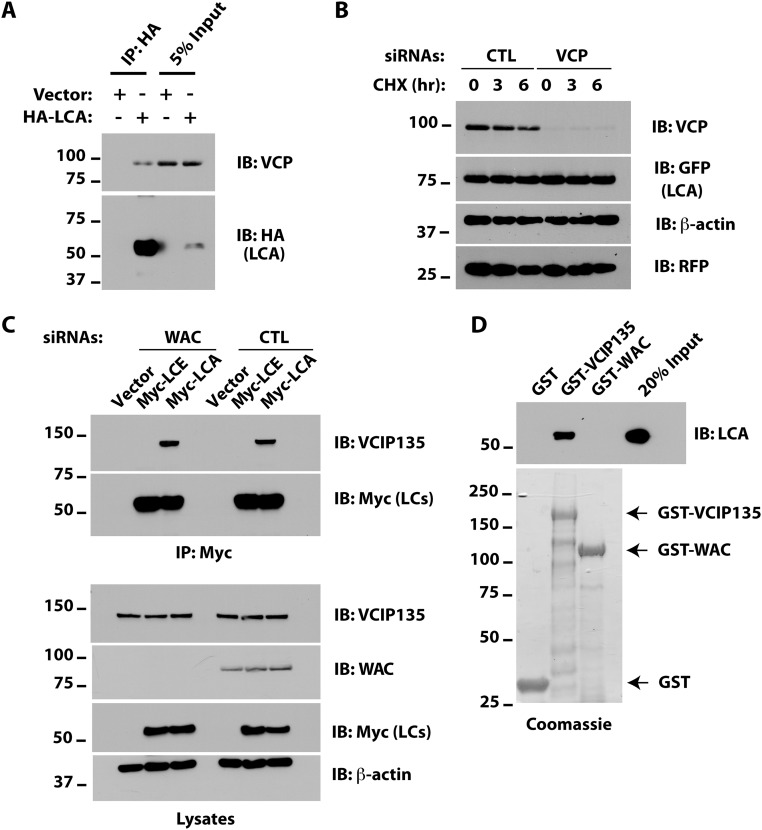
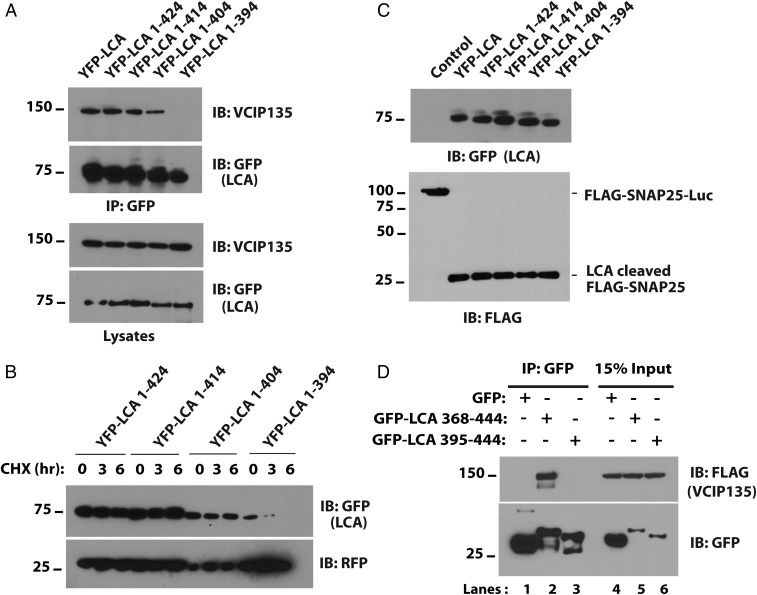
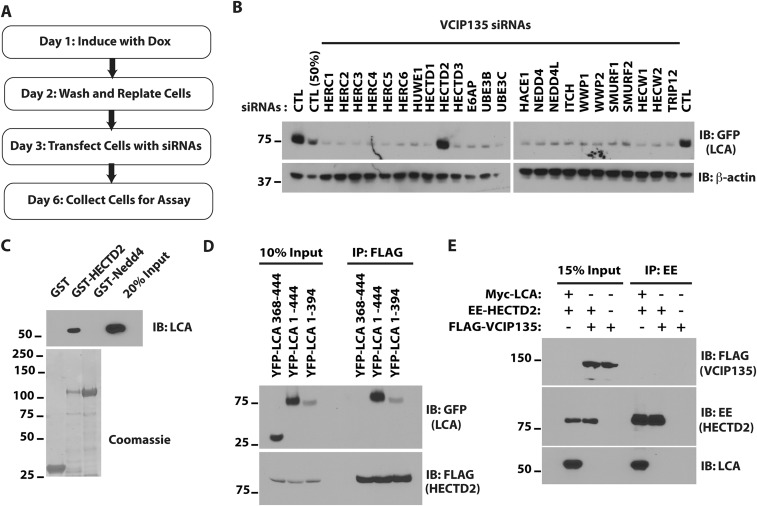
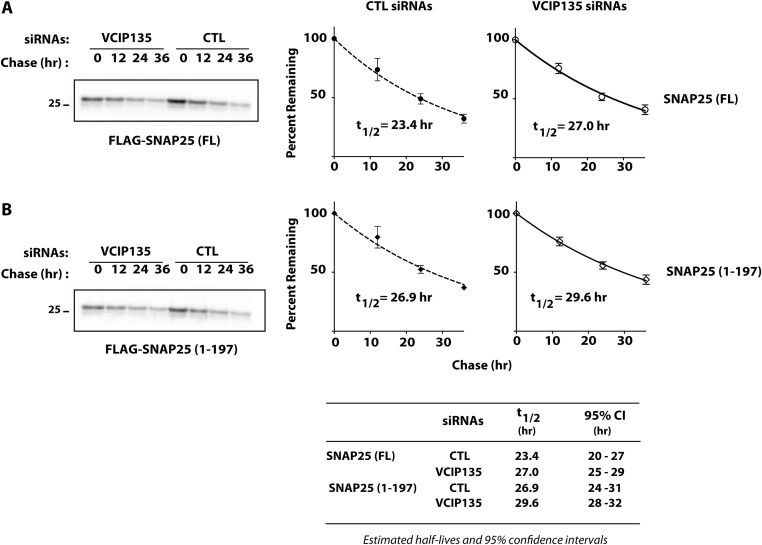
Botulism is characterized by flaccid paralysis, which can be caused by intoxication with any of the seven known serotypes of botulinum neurotoxin (BoNT), all of which disrupt synaptic transmission by endoproteolytic cleavage of SNARE proteins. BoNT serotype A (BoNT/A) has the most prolonged or persistent effects, which can last several months, and exerts its effects by specifically cleaving and inactivating SNAP25. A major factor contributing to the persistence of intoxication is the long half-life of the catalytic light chain, which remains enzymatically active months after entry into cells. Here we report that BoNT/A catalytic light chain binds to, and is a substrate for, the ubiquitin ligase HECTD2. However, the light chain evades proteasomal degradation by the dominant effect of a deubiquitinating enzyme, VCIP135/VCPIP1. This deubiquitinating enzyme binds BoNT/A light chain directly, with the two associating in cells through the C-terminal 77 amino acids of the light chain protease. The development of specific DUB inhibitors, together with inhibitors of BoNT/A proteolytic activity, may be useful for reducing the morbidity and public health costs associated with BoNT/A intoxication and could have potential biodefense implications.
Keywords: USP9X; motoneuron; synaptic transmission; synaptosomal-associated protein 25; toxin persistence.
Conflict of interest statement
Conflict of interest statement: G.A.O. has a financial interest in Synaptic Research LLC, which is developing therapeutics for BoNT intoxication, including the delivery of designer ubiquitin ligases.
Figures
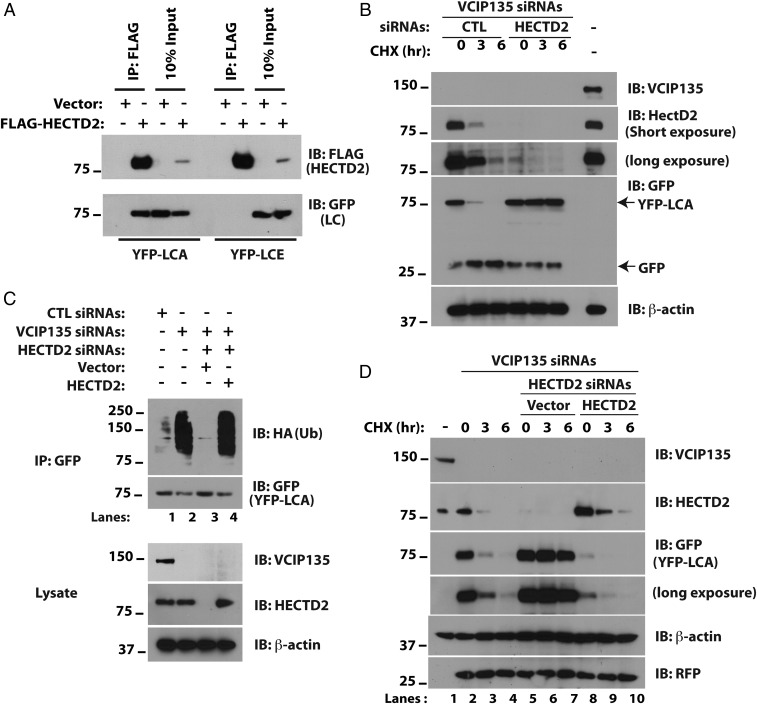
References
-
- Simpson LL. Identification of the characteristics that underlie botulinum toxin potency: Implications for designing novel drugs. Biochimie. 2000;82:943–953. - PubMed
-
- Habermann E, Dreyer F. Clostridial neurotoxins: Handling and action at the cellular and molecular level. Curr Top Microbiol Immunol. 1986;129:93–179. - PubMed
-
- Johnson EA, Montecucco C. Botulism. Handb Clin Neurol. 2008;91:333–368. - PubMed
-
- Schiavo G, Matteoli M, Montecucco C. Neurotoxins affecting neuroexocytosis. Physiol Rev. 2000;80:717–766. - PubMed
-
- Montecucco C, Schiavo G. Structure and function of tetanus and botulinum neurotoxins. Q Rev Biophys. 1995;28:423–472. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
