

Temperature variability is integrated by a spatially embedded decision-making center to break dormancy in Arabidopsis seeds
- PMID: 28584126
- PMCID: PMC5488954
- DOI: 10.1073/pnas.1704745114
Temperature variability is integrated by a spatially embedded decision-making center to break dormancy in Arabidopsis seeds
Abstract
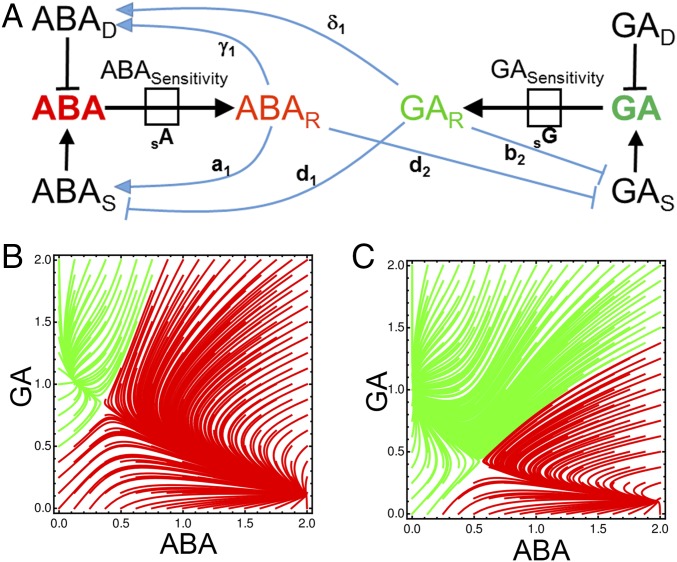
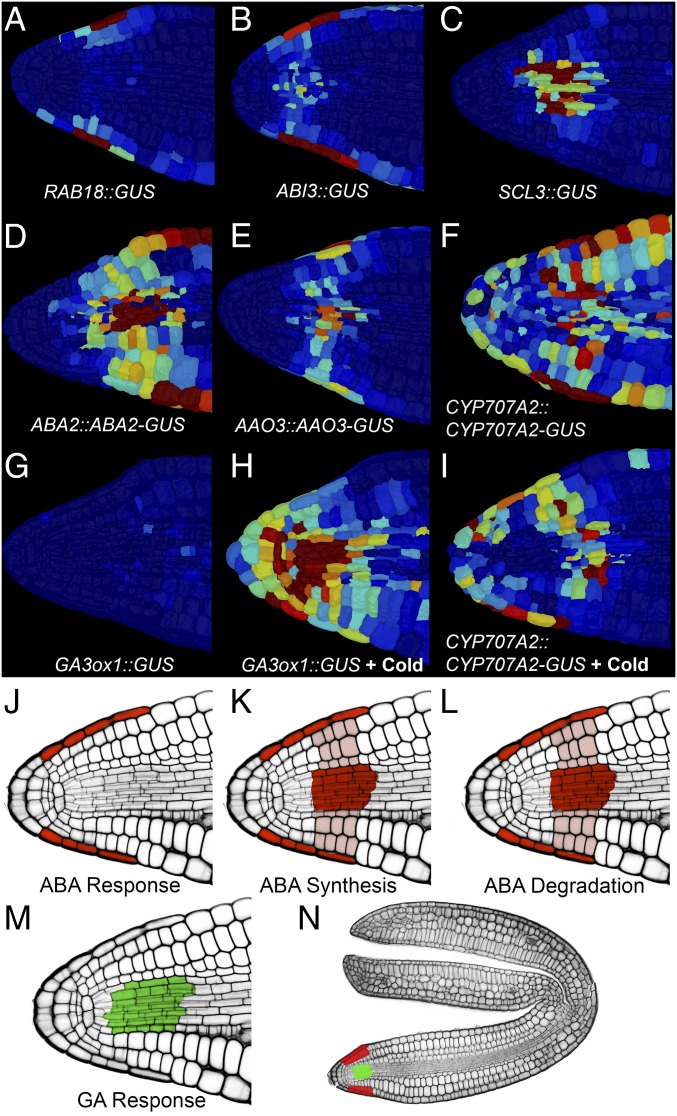
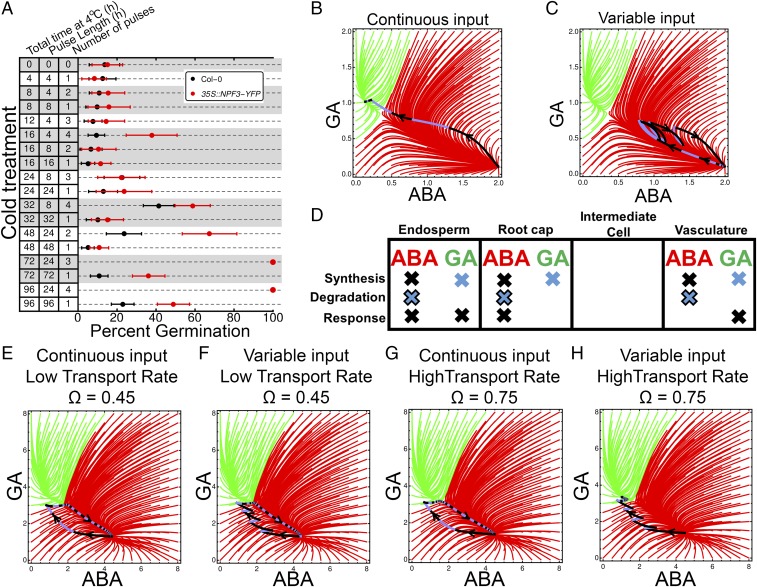
Plants perceive and integrate information from the environment to time critical transitions in their life cycle. Some mechanisms underlying this quantitative signal processing have been described, whereas others await discovery. Seeds have evolved a mechanism to integrate environmental information by regulating the abundance of the antagonistically acting hormones abscisic acid (ABA) and gibberellin (GA). Here, we show that hormone metabolic interactions and their feedbacks are sufficient to create a bistable developmental fate switch in Arabidopsis seeds. A digital single-cell atlas mapping the distribution of hormone metabolic and response components revealed their enrichment within the embryonic radicle, identifying the presence of a decision-making center within dormant seeds. The responses to both GA and ABA were found to occur within distinct cell types, suggesting cross-talk occurs at the level of hormone transport between these signaling centers. We describe theoretically, and demonstrate experimentally, that this spatial separation within the decision-making center is required to process variable temperature inputs from the environment to promote the breaking of dormancy. In contrast to other noise-filtering systems, including human neurons, the functional role of this spatial embedding is to leverage variability in temperature to transduce a fate-switching signal within this biological system. Fluctuating inputs therefore act as an instructive signal for seeds, enhancing the accuracy with which plants are established in ecosystems, and distributed computation within the radicle underlies this signal integration mechanism.
Keywords: distributed control; dormancy; seed; signal integration; variability.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Domagalska MA, Leyser O. Signal integration in the control of shoot branching. Nat Rev Mol Cell Biol. 2011;12:211–221. - PubMed
-
- Angel A, Song J, Dean C, Howard M. A Polycomb-based switch underlying quantitative epigenetic memory. Nature. 2011;476:105–108. - PubMed
-
- Finch-Savage WE, Leubner-Metzger G. Seed dormancy and the control of germination. New Phytol. 2006;171:501–523. - PubMed
-
- Finch-Savage WE, Bassel GW. Seed vigour and crop establishment: Extending performance beyond adaptation. J Exp Bot. 2016;67:567–591. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
