

Card9-dependent IL-1β regulates IL-22 production from group 3 innate lymphoid cells and promotes colitis-associated cancer
- PMID: 28586167
- PMCID: PMC5600091
- DOI: 10.1002/eji.201646765
Card9-dependent IL-1β regulates IL-22 production from group 3 innate lymphoid cells and promotes colitis-associated cancer
Abstract
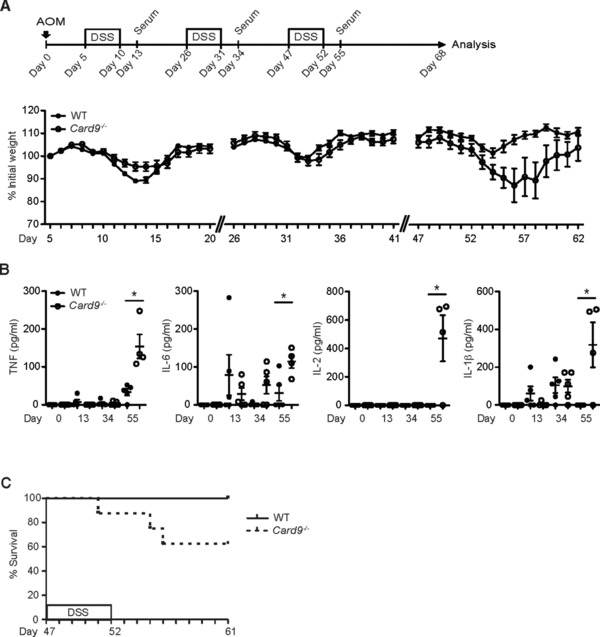
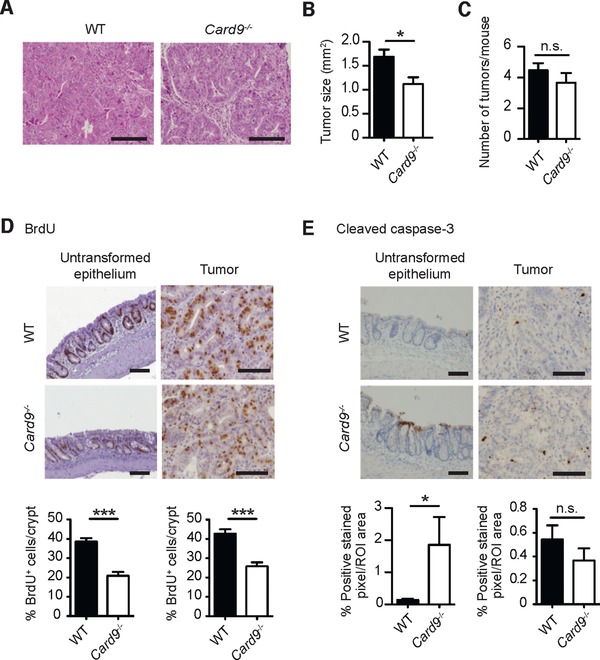
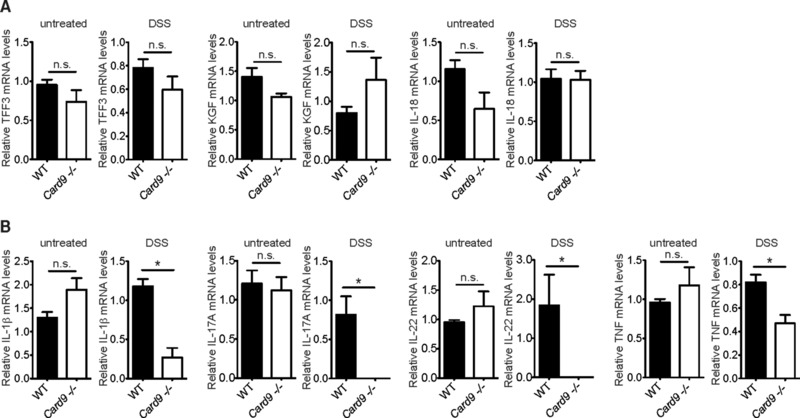
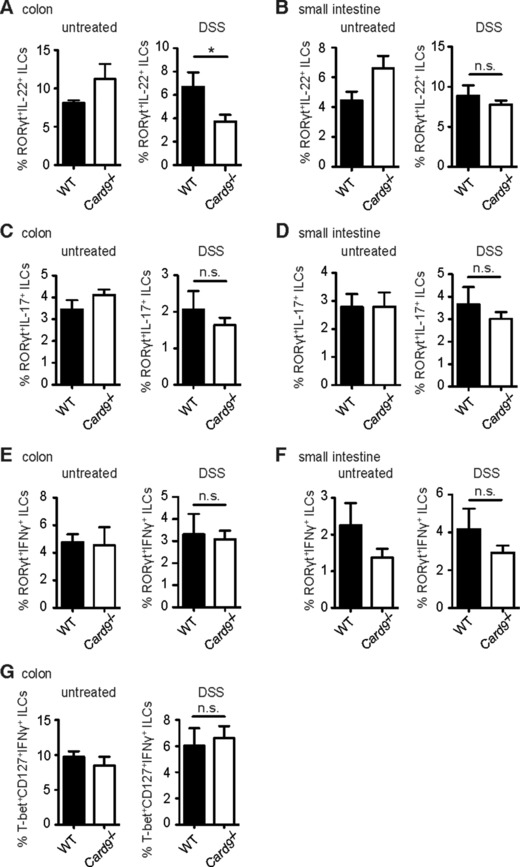
Inflammatory bowel diseases (IBD) are key risk factors for the development of colorectal cancer, but the mechanisms that link intestinal inflammation with carcinogenesis are insufficiently understood. Card9 is a myeloid cell-specific signaling protein that regulates inflammatory responses downstream of various pattern recognition receptors and which cooperates with the inflammasomes for IL-1β production. Because polymorphisms in Card9 were recurrently associated with human IBD, we investigated the function of Card9 in a colitis-associated cancer (CAC) model. Card9-/- mice develop smaller, less proliferative and less dysplastic tumors compared to their littermates and in the regenerating mucosa we detected dramatically impaired IL-1β generation and defective IL-1β controlled IL-22 production from group 3 innate lymphoid cells. Consistent with the key role of immune-derived IL-22 in activating STAT3 signaling during normal and pathological intestinal epithelial cell (IEC) proliferation, Card9-/- mice also exhibit impaired tumor cell intrinsic STAT3 activation. Our results imply a Card9-controlled, ILC3-mediated mechanism regulating healthy and malignant IEC proliferation and demonstrates a role of Card9-mediated innate immunity in inflammation-associated carcinogenesis.
Keywords: Card9; Colitis-associated-cancer; Innate lymphoid cells; Interleukin-1β; Interleukin-22.
© 2017 The Authors. European Journal of Immunology published by WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim.
Figures
References
-
- Beaugerie, L. and Itzkowitz, S. H. , Cancers complicating inflammatory bowel disease. N. Engl. J. Med. 2015. 372: 1441–1452. - PubMed
-
- Zhernakova, A. , Festen, E. M. , Franke, L. , Trynka, G. , van Diemen, C. C. , Monsuur, A. J. , Bevova, M. et al., Genetic analysis of innate immunity in Crohn's disease and ulcerative colitis identifies two susceptibility loci harboring CARD9 and IL18RAP. Am. J. Hum. Genet. 2008. 82: 1202–1210. - PMC - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
