

Molecular Basis for K63-Linked Ubiquitination Processes in Double-Strand DNA Break Repair: A Focus on Kinetics and Dynamics
- PMID: 28587922
- PMCID: PMC7507754
- DOI: 10.1016/j.jmb.2017.05.029
Molecular Basis for K63-Linked Ubiquitination Processes in Double-Strand DNA Break Repair: A Focus on Kinetics and Dynamics
Abstract
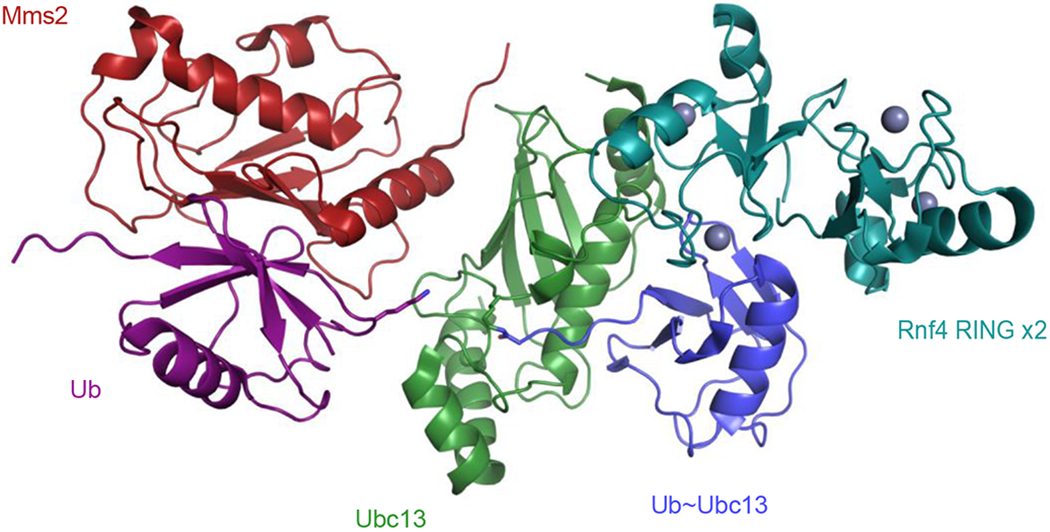
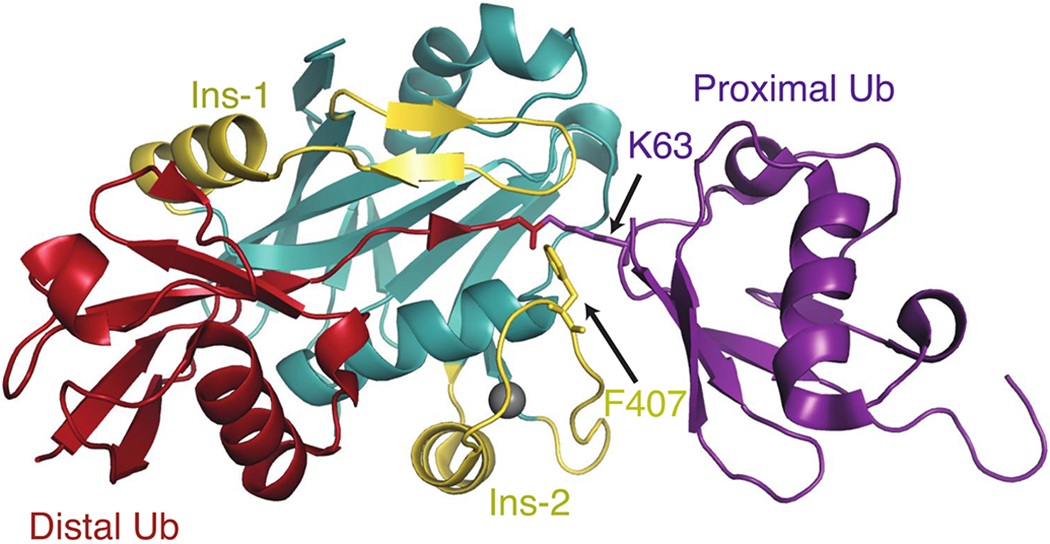
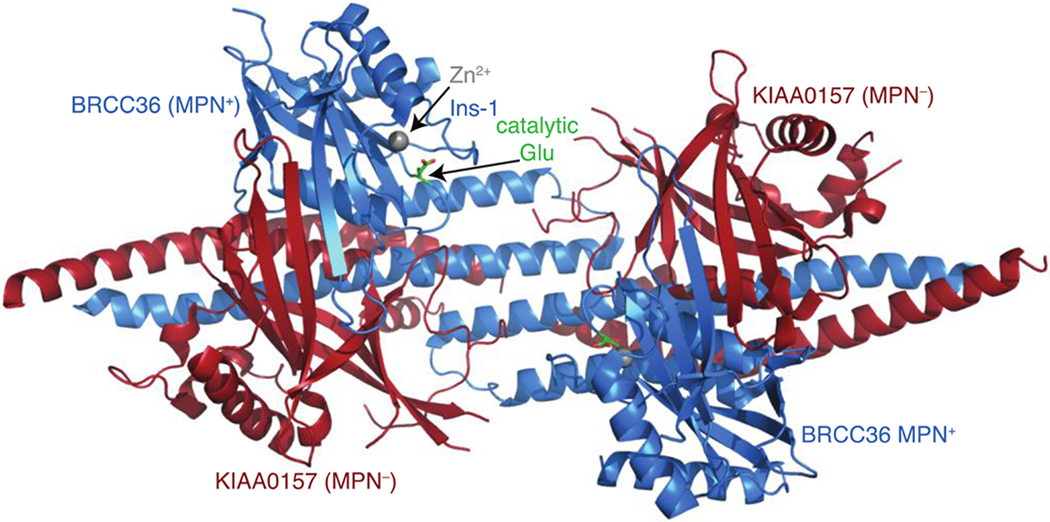
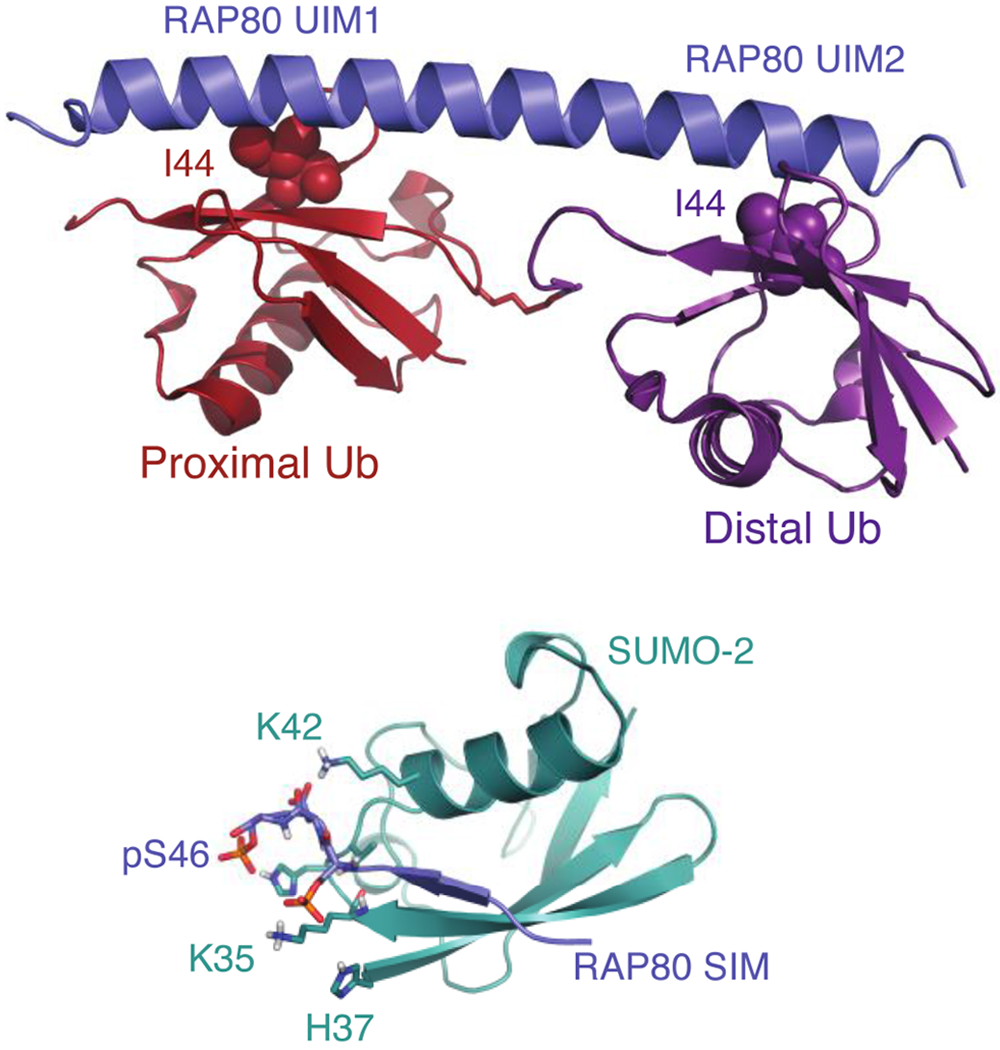
Cells are exposed to thousands of DNA damage events on a daily basis. This damage must be repaired to preserve genetic information and prevent development of disease. The most deleterious damage is a double-strand break (DSB), which is detected and repaired by mechanisms known as non-homologous end-joining (NHEJ) and homologous recombination (HR), which are components of the DNA damage response system. NHEJ is an error-prone first line of defense, whereas HR invokes error-free repair and is the focus of this review. The functions of the protein components of HR-driven DNA repair are regulated by the coordinated action of post-translational modifications including lysine acetylation, phosphorylation, ubiquitination, and SUMOylation. The latter two mechanisms are fundamental for recognition of DSBs and reorganizing chromatin to facilitate repair. We focus on the structures and molecular mechanisms for the protein components underlying synthesis, recognition, and cleavage of K63-linked ubiquitin chains, which are abundant at damage sites and obligatory for DSB repair. The forward flux of the K63-linked ubiquitination cascade is driven by the combined activity of E1 enzyme, the heterodimeric E2 Mms2-Ubc13, and its cognate E3 ligases RNF8 and RNF168, which is balanced through the binding and cleavage of chains by the deubiquitinase BRCC36, and the proteasome, and through the binding of chains by recognition modules on repair proteins such as RAP80. We highlight a number of aspects regarding our current understanding for the role of kinetics and dynamics in determining the function of the enzymes and chain recognition modules that drive K63 ubiquitination.
Keywords: DNA damage response; enzyme kinetics; protein dynamics; protein–protein-interactions; ubiquitination.
Copyright © 2017 Elsevier Ltd. All rights reserved.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
