

Effect of water flow and chemical environment on microbiota growth and composition in the human colon
- PMID: 28588144
- PMCID: PMC5488924
- DOI: 10.1073/pnas.1619598114
Effect of water flow and chemical environment on microbiota growth and composition in the human colon
Abstract
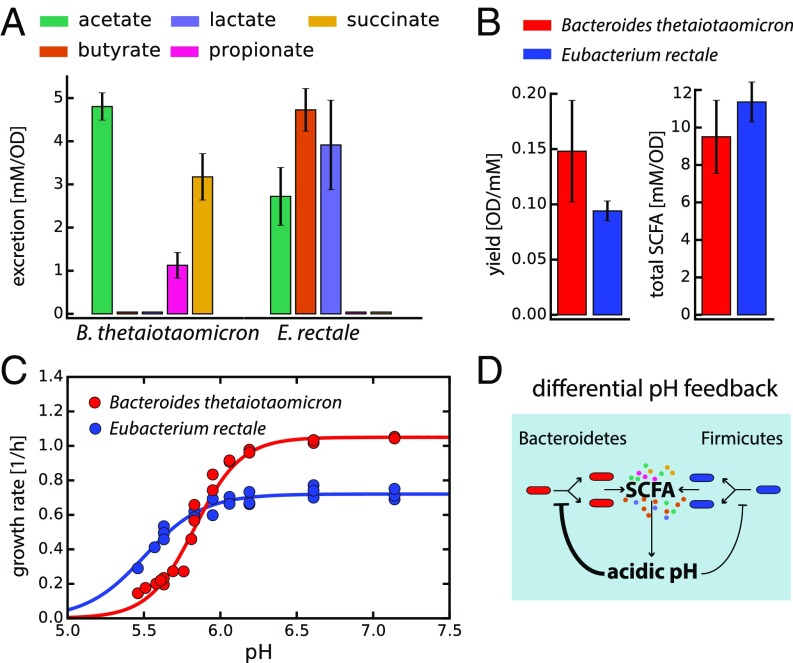
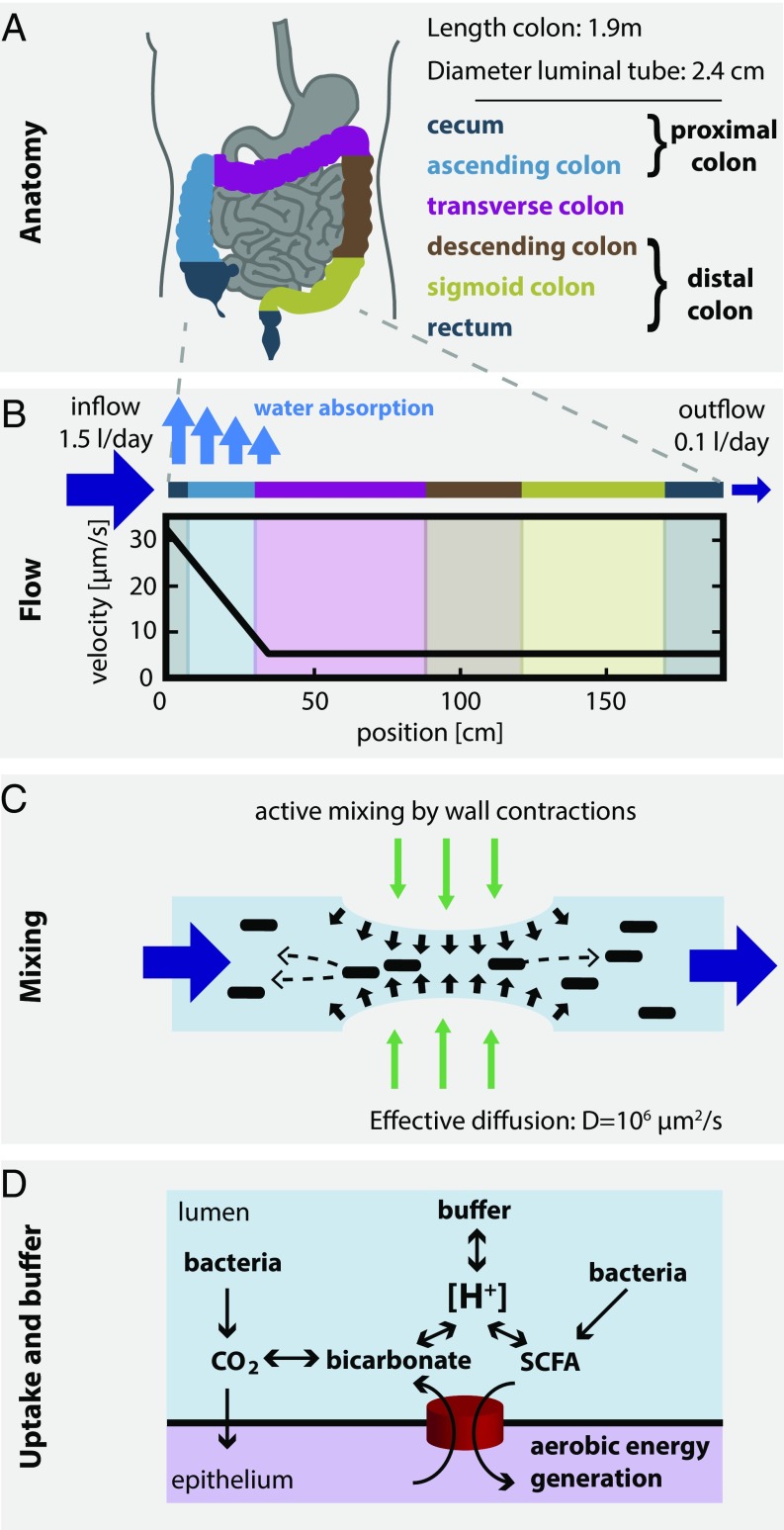
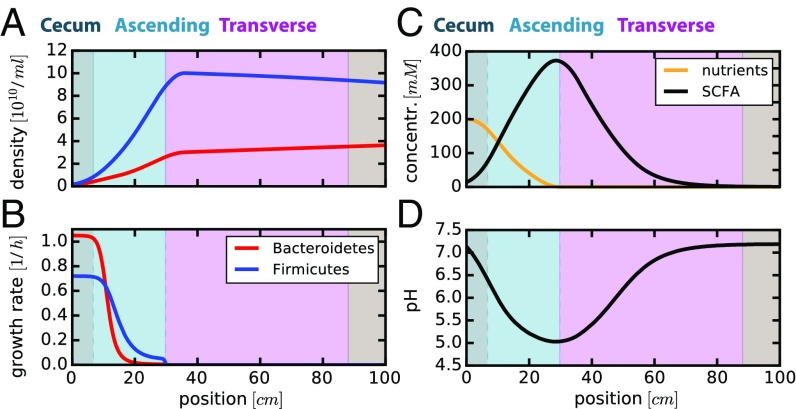
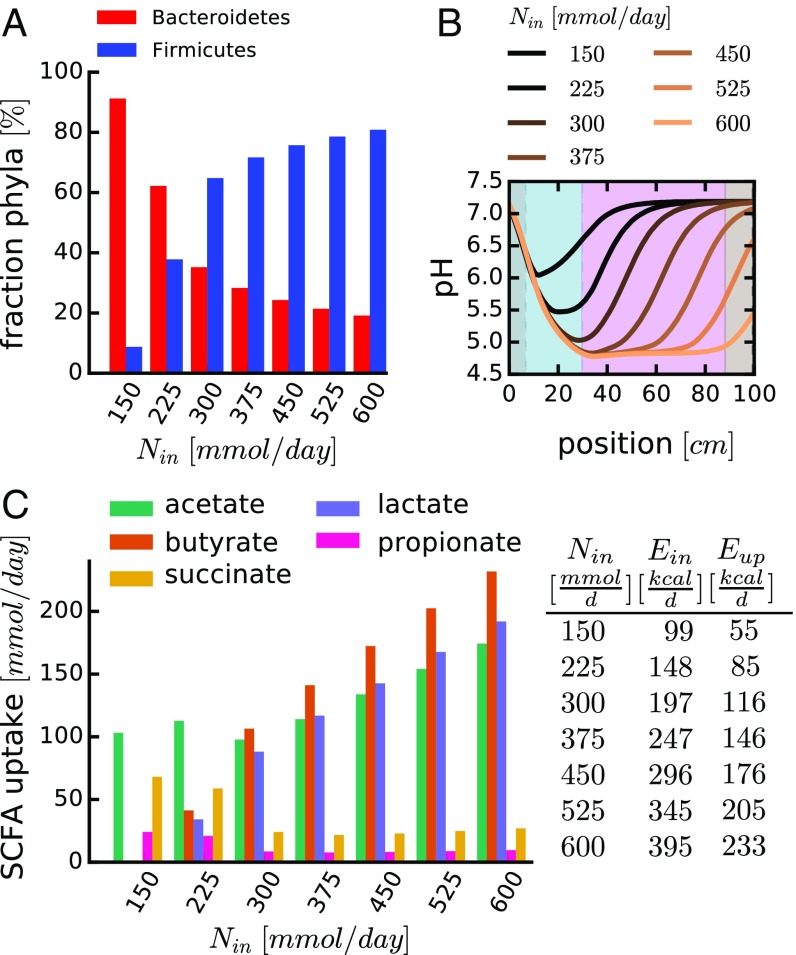
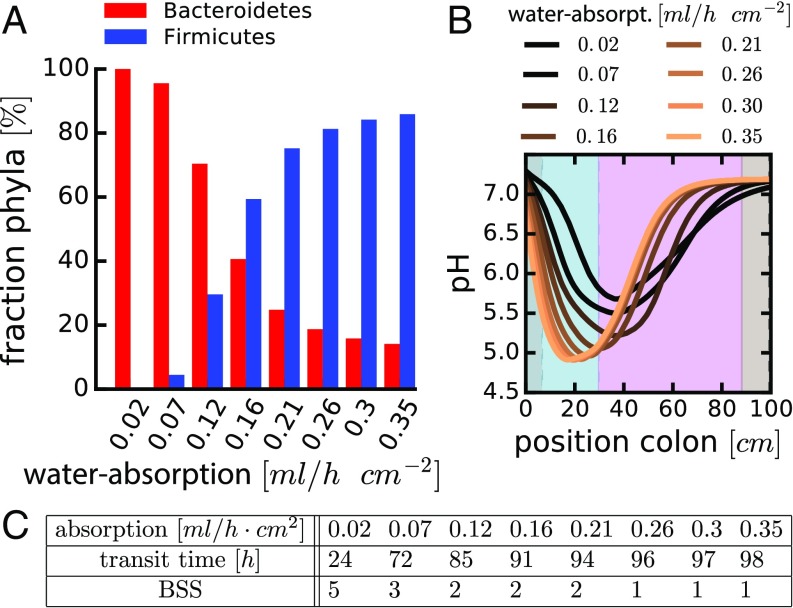
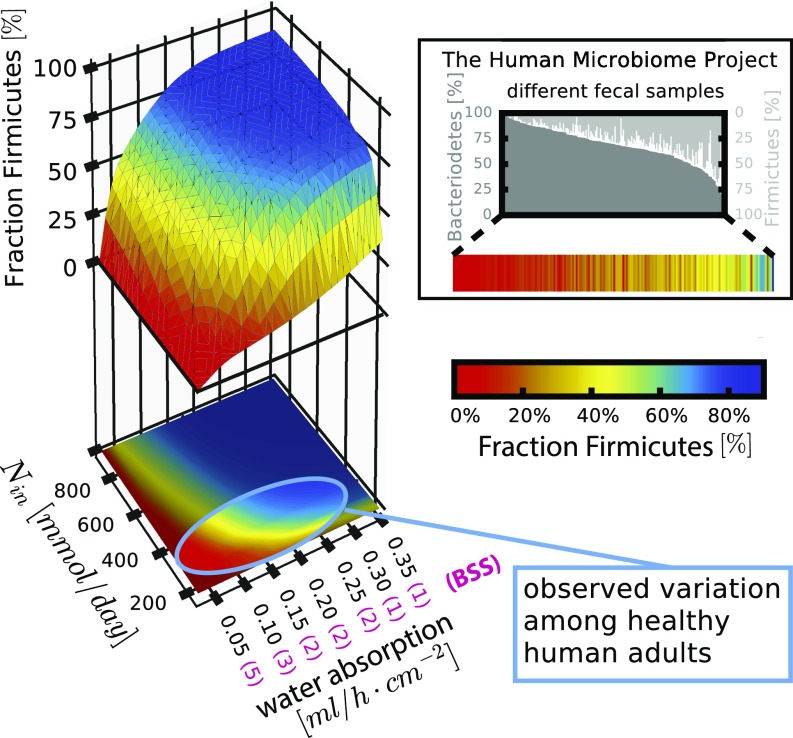
The human gut harbors a dynamic microbial community whose composition bears great importance for the health of the host. Here, we investigate how colonic physiology impacts bacterial growth, which ultimately dictates microbiota composition. Combining measurements of bacterial physiology with analysis of published data on human physiology into a quantitative, comprehensive modeling framework, we show how water flow in the colon, in concert with other physiological factors, determine the abundances of the major bacterial phyla. Mechanistically, our model shows that local pH values in the lumen, which differentially affect the growth of different bacteria, drive changes in microbiota composition. It identifies key factors influencing the delicate regulation of colonic pH, including epithelial water absorption, nutrient inflow, and luminal buffering capacity, and generates testable predictions on their effects. Our findings show that a predictive and mechanistic understanding of microbial ecology in the gut is possible. Such predictive understanding is needed for the rational design of intervention strategies to actively control the microbiota.
Keywords: colon physiology; colonic pH; gut microbiota; stool consistency; water absorption.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
