

Melon13-lipoxygenase CmLOX18 may be involved in C6 volatiles biosynthesis in fruit
- PMID: 28588227
- PMCID: PMC5460189
- DOI: 10.1038/s41598-017-02559-6
Melon13-lipoxygenase CmLOX18 may be involved in C6 volatiles biosynthesis in fruit
Abstract
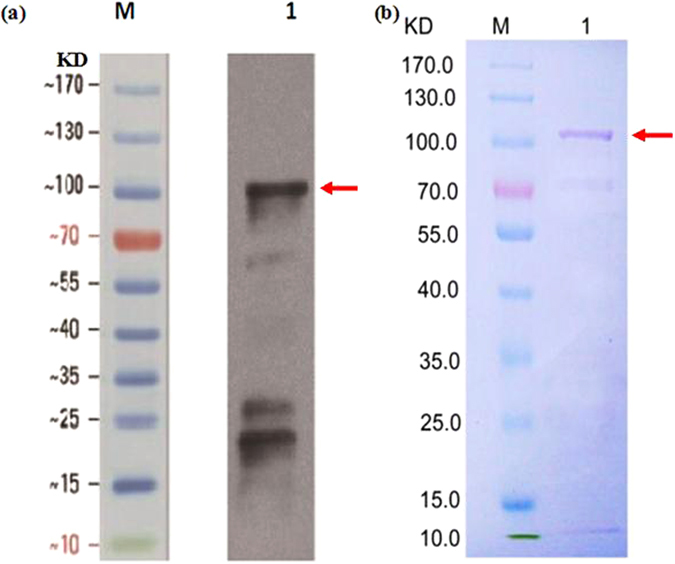
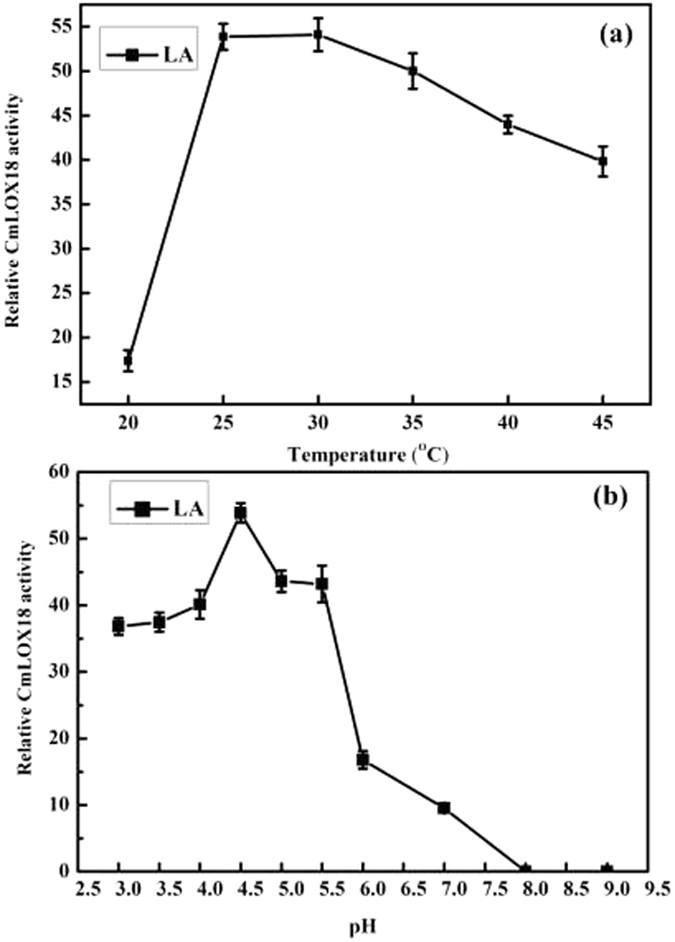
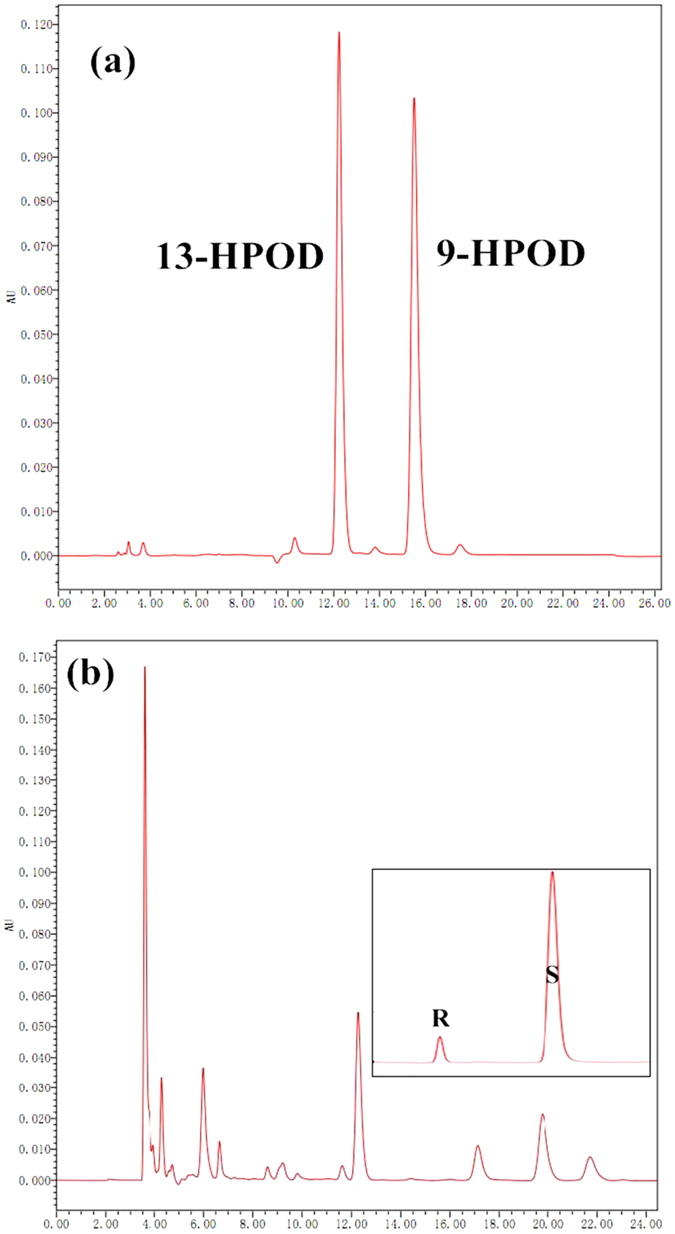
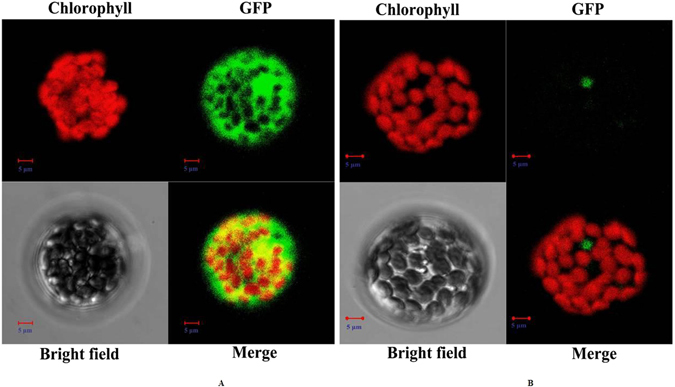
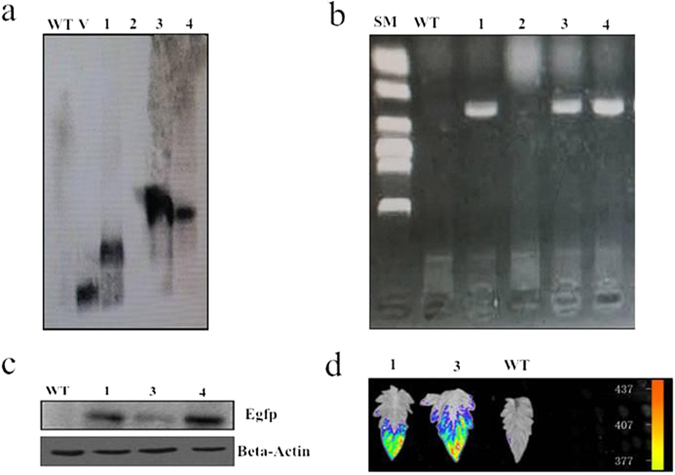
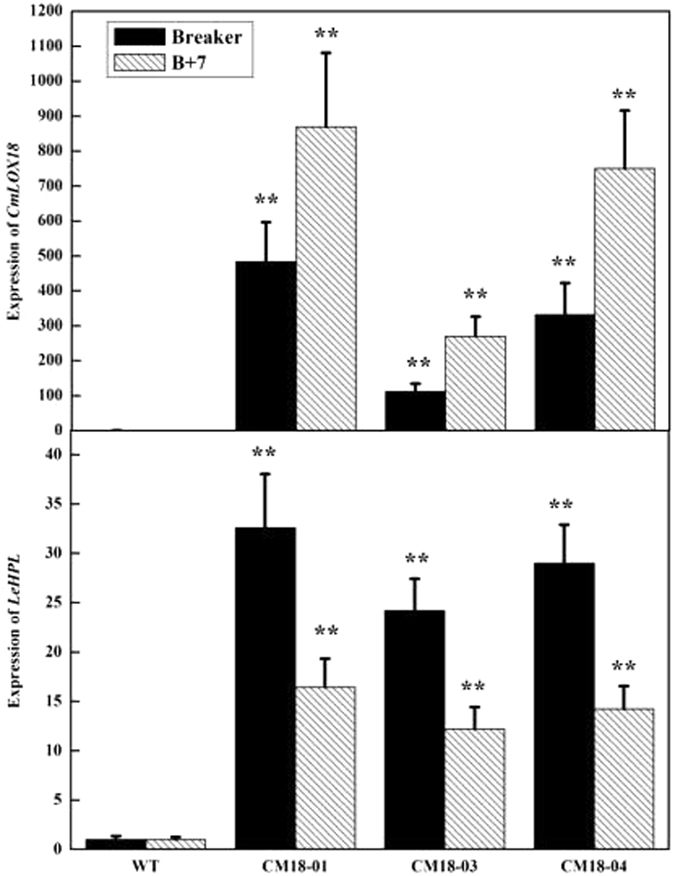
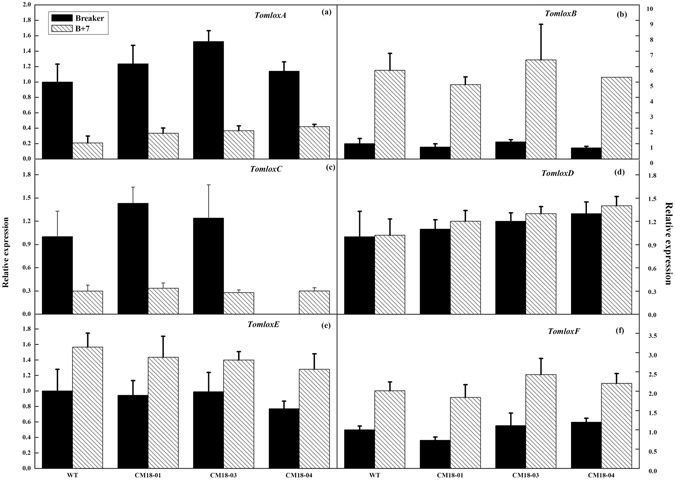
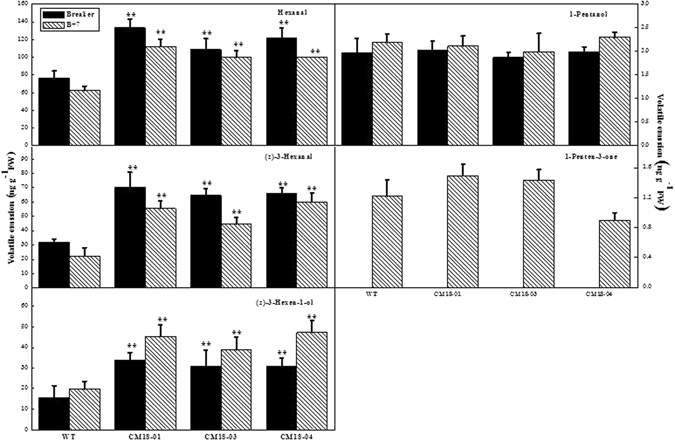
To better understand the function role of the melon CmLOX18 gene in the biosynthesis of C6 volatiles during fruit ripening, we biochemically characterized CmLOX18 and identified its subcellular localization in transgenic tomato plants. Heterologous expression in yeast cells showed that the molecular weight of the CmLOX18 protein was identical to that predicted, and that this enzyme possesseed lipoxygenase activity. Linoleic acid was demonstrated to be the preferred substrate for the purified recombinant CmLOX18 protein, which exhibited optimal catalytic activity at pH 4.5 and 30 °C. Chromatogram analysis of the reaction product indicated that the CmLOX18 protein exhibited positional specificity, as evidenced by its release of only a C-13 oxidized product. Subcellular localization analysis by transient expression in Arabidopsis protoplasts showed that CmLOX18 was localized to non-chloroplast organelles. When the CmLOX18 gene was transgenically expressed in tomato via Agrobacterium tumefaciens-mediated transformation, it was shown to enhance expression levels of the tomato hydroperoxide lyase gene LeHPL, whereas the expression levels of six TomLox genes were little changed. Furthermore, transgenic tomato fruits exhibited increases in the content of the C6 volatiles, namely hexanal, (Z)-3-hexanal, and (Z)-3-hexen-1-ol, indicating that CmLOX18 probably plays an important role in the synthesis of C6 compounds in fruits.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures
Similar articles
-
A 13-lipoxygenase, TomloxC, is essential for synthesis of C5 flavour volatiles in tomato.J Exp Bot. 2014 Feb;65(2):419-28. doi: 10.1093/jxb/ert382. Epub 2014 Jan 22. J Exp Bot. 2014. PMID: 24453226 Free PMC article.
-
Simultaneous silencing of five lipoxygenase genes increases the contents of α-linolenic and linoleic acids in tomato (Solanum lycopersicum L.) fruits.J Agric Food Chem. 2014 Dec 10;62(49):11988-93. doi: 10.1021/jf503801u. Epub 2014 Dec 1. J Agric Food Chem. 2014. PMID: 25418937
-
Contribution of abscisic acid to aromatic volatiles in cherry tomato (Solanum lycopersicum L.) fruit during postharvest ripening.Plant Physiol Biochem. 2018 Sep;130:205-214. doi: 10.1016/j.plaphy.2018.06.039. Epub 2018 Jun 30. Plant Physiol Biochem. 2018. PMID: 29990773
-
The genetic basis of fruit morphology in horticultural crops: lessons from tomato and melon.J Exp Bot. 2014 Aug;65(16):4625-37. doi: 10.1093/jxb/eru017. Epub 2014 Feb 11. J Exp Bot. 2014. PMID: 24520021 Review.
-
Fatty acid hydroperoxide lyase: a plant cytochrome p450 enzyme involved in wound healing and pest resistance.Chembiochem. 2001 Aug 3;2(7-8):494-504. doi: 10.1002/1439-7633(20010803)2:7/8<494::AID-CBIC494>3.0.CO;2-1. Chembiochem. 2001. PMID: 11828481 Review.
Cited by
-
The effects of different rootstocks on aroma components, activities and genes expression of aroma-related enzymes in oriental melon fruit.PeerJ. 2024 Jan 5;12:e16704. doi: 10.7717/peerj.16704. eCollection 2024. PeerJ. 2024. PMID: 38192601 Free PMC article.
-
Molecular, in silico and expression studies on lipoxygenases (LOXs) in potato (Solanum tuberosum L.).3 Biotech. 2023 Dec;13(12):419. doi: 10.1007/s13205-023-03839-x. Epub 2023 Nov 28. 3 Biotech. 2023. PMID: 38037658 Free PMC article.
-
Postharvest chilling diminishes melon flavor via effects on volatile acetate ester biosynthesis.Front Plant Sci. 2023 Jan 6;13:1067680. doi: 10.3389/fpls.2022.1067680. eCollection 2022. Front Plant Sci. 2023. PMID: 36684781 Free PMC article.
-
Genome-Wide Identification and Expression Analysis of NRAMP Family Genes in Soybean (Glycine Max L.).Front Plant Sci. 2017 Aug 18;8:1436. doi: 10.3389/fpls.2017.01436. eCollection 2017. Front Plant Sci. 2017. PMID: 28868061 Free PMC article.
-
From Central to Specialized Metabolism: An Overview of Some Secondary Compounds Derived From the Primary Metabolism for Their Role in Conferring Nutritional and Organoleptic Characteristics to Fruit.Front Plant Sci. 2019 Jun 28;10:835. doi: 10.3389/fpls.2019.00835. eCollection 2019. Front Plant Sci. 2019. PMID: 31316537 Free PMC article. Review.
References
-
- Defilippi BG, Manríquez D, Luengwilai K, González-Agüero M. Chapter 1 Aroma Volatiles: Biosynthesis and Mechanisms of Modulation During Fruit Ripening. Adv. Bot. Res. 2009;50:1–37. doi: 10.1016/S0065-2296(08)00801-X. - DOI
-
- Dirinck P, De Pooter H, Schamp N. Aroma development in ripening fruits. In: Flavor chemistry: trends and developments. Teranishi R and Buttery R (editors) American Chemical Society (ACS) Symposium series. 1989;388:23–34.
-
- Song J, Forney CF. Flavour volatile production and regulation in fruit. Can J Plant Sc i/Rev Can Phytotechi. 2008;88:537–550. doi: 10.4141/CJPS07170. - DOI
-
- Baldwin EA, Scott JW, Shewmaker CK, Schuch W. Flavor trivia and tomato aroma: biochemistry and possible mechanisms for control of important aroma components. Hort. sci. 2000;5:1013–1022.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
