

Quantum effects in biology: golden rule in enzymes, olfaction, photosynthesis and magnetodetection
- PMID: 28588400
- PMCID: PMC5454345
- DOI: 10.1098/rspa.2016.0822
Quantum effects in biology: golden rule in enzymes, olfaction, photosynthesis and magnetodetection
Abstract
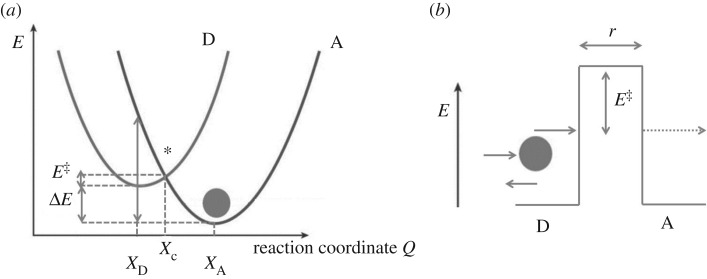
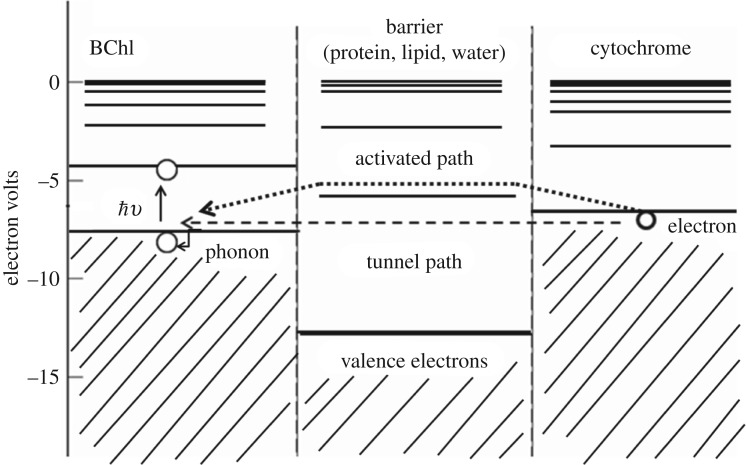
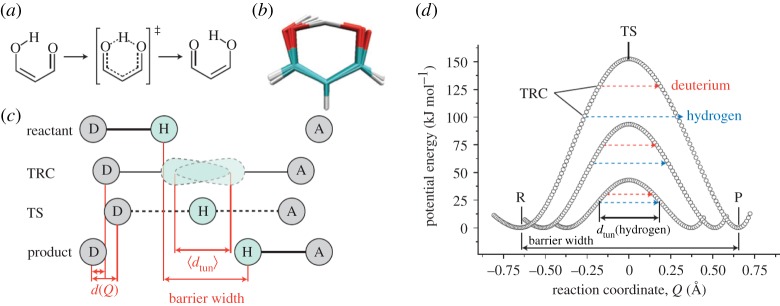
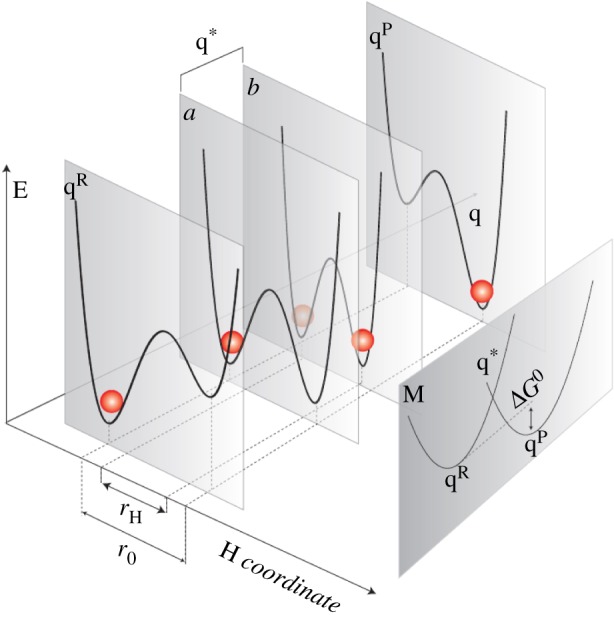
Despite certain quantum concepts, such as superposition states, entanglement, 'spooky action at a distance' and tunnelling through insulating walls, being somewhat counterintuitive, they are no doubt extremely useful constructs in theoretical and experimental physics. More uncertain, however, is whether or not these concepts are fundamental to biology and living processes. Of course, at the fundamental level all things are quantum, because all things are built from the quantized states and rules that govern atoms. But when does the quantum mechanical toolkit become the best tool for the job? This review looks at four areas of 'quantum effects in biology'. These are biosystems that are very diverse in detail but possess some commonality. They are all (i) effects in biology: rates of a signal (or information) that can be calculated from a form of the 'golden rule' and (ii) they are all protein-pigment (or ligand) complex systems. It is shown, beginning with the rate equation, that all these systems may contain some degree of quantumeffect, and where experimental evidence is available, it is explored to determine how the quantum analysis aids in understanding of the process.
Keywords: enzymes; magnetodetection; olfaction; photosynthesis; quantum.
Conflict of interest statement
I declare I have no competing interests.
Figures
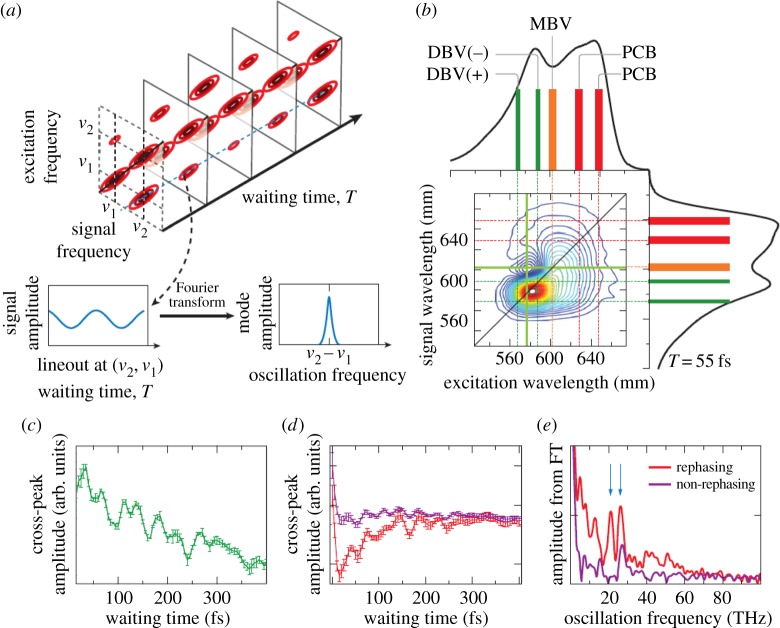
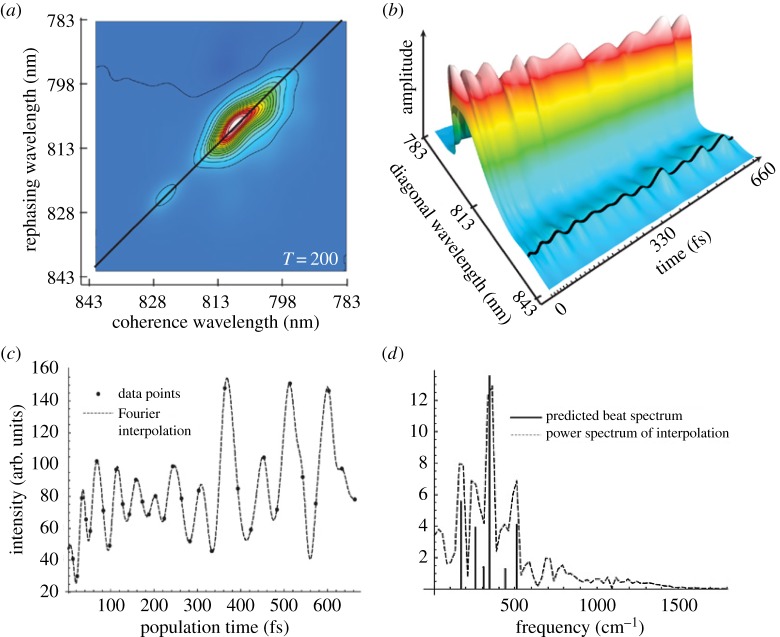
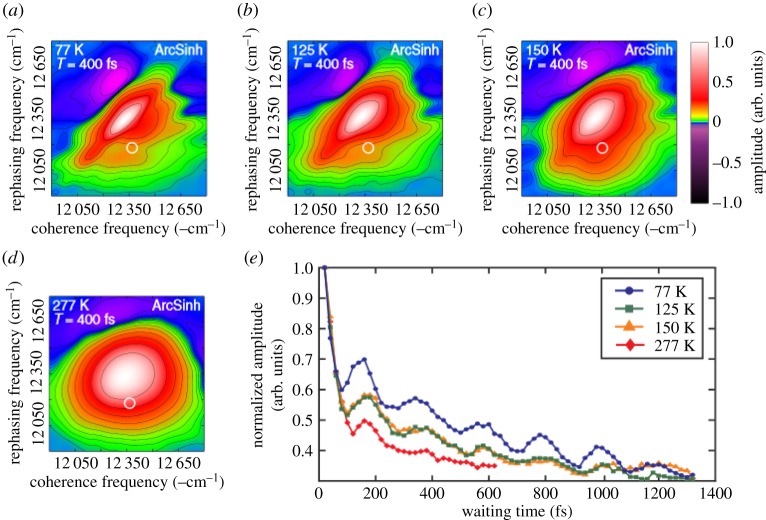
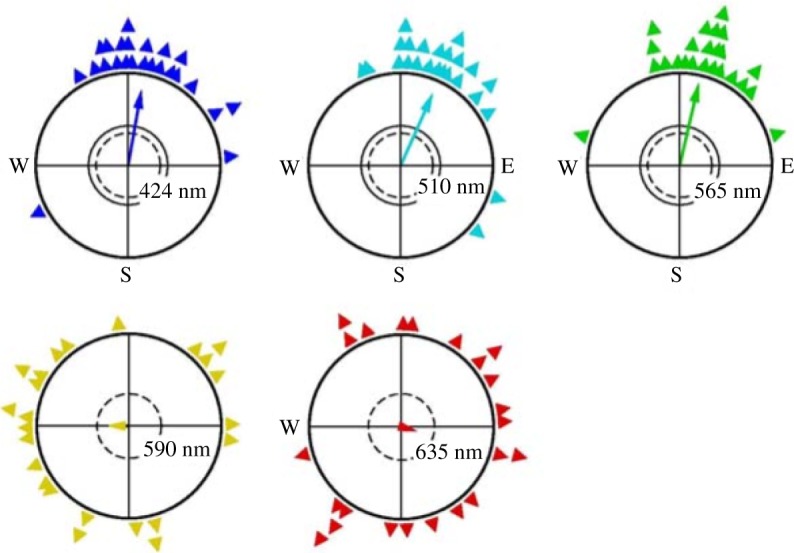
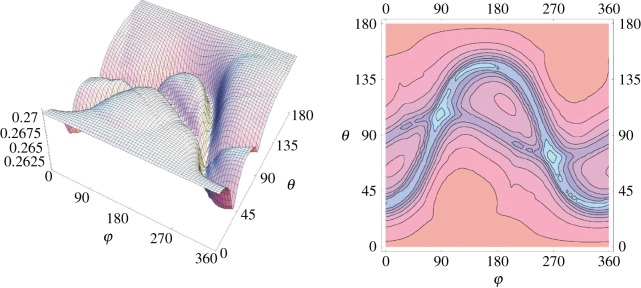
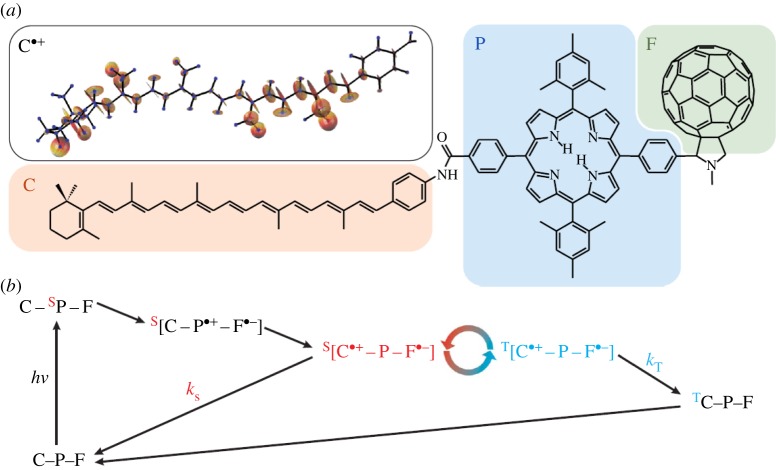
References
-
- De Vault D. 1984. Quantum-mechanical tunneling in biological systems, vol. 2 Cambridge, UK: Cambridge University Press.
-
- Klinman JP, Kohen A. 2013. Hydrogen tunneling links protein dynamics to enzyme catalysis. Annu. Rev. Biochem. 82, 471–496. (doi:10.1146/annurev-biochem-051710-133623) - DOI - PMC - PubMed
-
- Eyring H. 1935. The activated complex and the absolute rate of chemical reactions. Chem. Rev. 17, 65–77. (doi:10.1021/cr60056a006) - DOI
-
- Gray HB, Winkler JR. 2005. Long-range electron transfer. Proc. Natl Acad. Sci. USA 102, 3534–3539. (doi:10.1073/pnas.0408029102) - DOI - PMC - PubMed
-
- Oppenheimer JR. 1928. Three notes on the quantum theory of aperiodic effects. Phys. Rev. 31, 66–81. (doi:10.1103/PhysRev.31.66) - DOI
Publication types
LinkOut - more resources
Full Text Sources
Other Literature Sources