

Forty Years of Sodium Channels: Structure, Function, Pharmacology, and Epilepsy
- PMID: 28589518
- PMCID: PMC5693772
- DOI: 10.1007/s11064-017-2314-9
Forty Years of Sodium Channels: Structure, Function, Pharmacology, and Epilepsy
Abstract
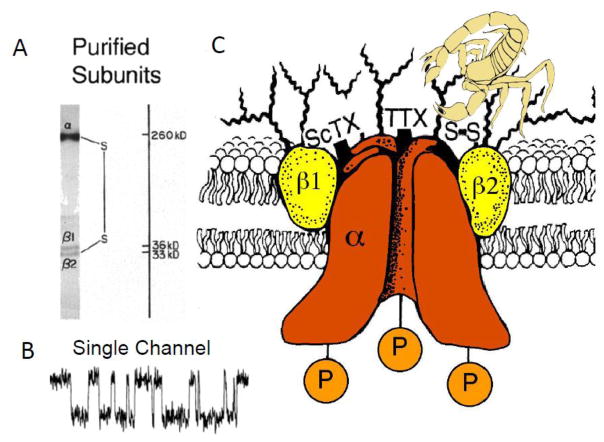
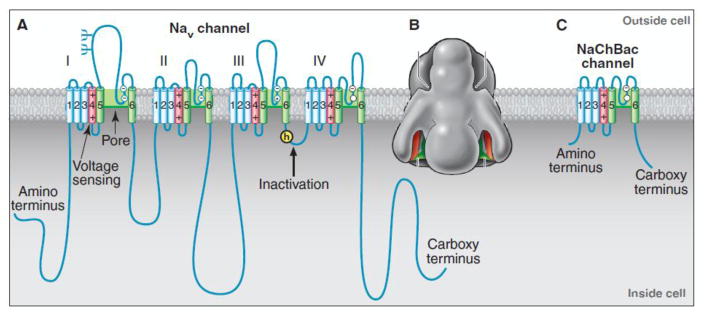
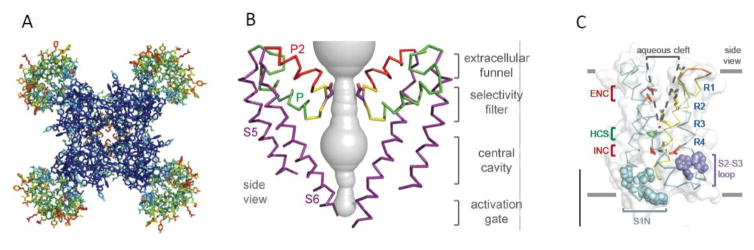
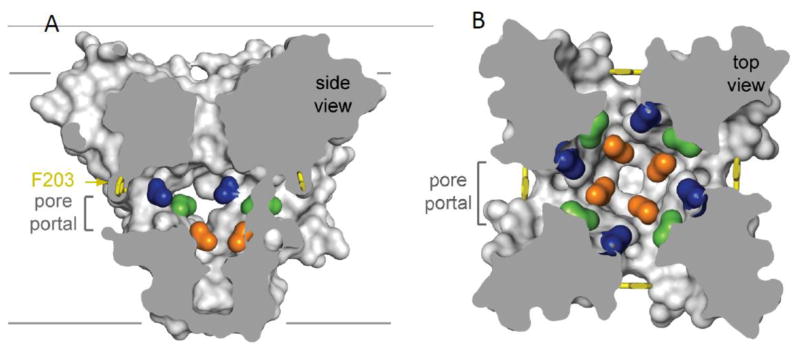
Voltage-gated sodium channels initiate action potentials in brain neurons. In the 1970s, much was known about the function of sodium channels from measurements of ionic currents using the voltage clamp method, but there was no information about the sodium channel molecules themselves. As a postdoctoral fellow and staff scientist at the National Institutes of Health, I developed neurotoxins as molecular probes of sodium channels in cultured neuroblastoma cells. During those years, Bruce Ransom and I crossed paths as members of the laboratories of Marshall Nirenberg and Philip Nelson and shared insights about sodium channels in neuroblastoma cells from my work and electrical excitability and synaptic transmission in cultured spinal cord neurons from Bruce's pioneering electrophysiological studies. When I established my laboratory at the University of Washington in 1977, my colleagues and I used those neurotoxins to identify the protein subunits of sodium channels, purify them, and reconstitute their ion conductance activity in pure form. Subsequent studies identified the molecular basis for the main functions of sodium channels-voltage-dependent activation, rapid and selective ion conductance, and fast inactivation. Bruce Ransom and I re-connected in the 1990s, as ski buddies at the Winter Conference on Brain Research and as faculty colleagues at the University of Washington when Bruce became our founding Chair of Neurology and provided visionary leadership of that department. In the past decade my work on sodium channels has evolved into structural biology. Molecular modeling and X-ray crystallographic studies have given new views of sodium channel function at atomic resolution. Sodium channels are also the molecular targets for genetic diseases, including Dravet Syndrome, an intractable pediatric epilepsy disorder with major co-morbidities of cognitive deficit, autistic-like behaviors, and premature death that is caused by loss-of-function mutations in the brain sodium channel NaV1.1. Our work on a mouse genetic model of this disease has shown that its multi-faceted pathophysiology and co-morbidities derive from selective loss of electrical excitability and action potential firing in GABAergic inhibitory neurons, which disinhibits neural circuits throughout the brain and leads directly to the epilepsy, premature death and complex co-morbidities of this disease. It has been rewarding for me to use our developing knowledge of sodium channels to help understand the pathophysiology and to suggest potential therapeutic approaches for this devastating childhood disease.
Keywords: Epilepsy; Ion channel structure; Local anesthetics; Sodium channel.
Figures
References
-
- Ransom BR, Barker JL, Nelson PG. Two mechanisms for poststimulus hyperpolarisations in cultured mammalian neurones. Nature. 1975;256:424–425. - PubMed
-
- Ransom BR, Bullock PN, Nelson PG. Mouse spinal cord in cell culture. III. Neuronal chemosensitivity and its relationship to synaptic activity. J Neurophysiol. 1977;40:1163–1177. - PubMed
-
- Ransom BR, Christian CN, Bullock PN, Nelson PG. Mouse spinal cord in cell culture. II. Synaptic activity and circuit behavior. J Neurophysiol. 1977;40:1151–1162. - PubMed
-
- Nelson PG, Ransom BR, Henkart M, Bullock PN. Mouse spinal cord in cell culture. IV. Modulation of inhibitory synaptic function. J Neurophysiol. 1977;40:1178–1187. - PubMed
-
- Ransom BR, Neale E, Henkart M, Bullock PN, Nelson PG. Mouse spinal cord in cell culture. I. Morphology and intrinsic neuronal electrophysiologic properties. J Neurophysiol. 1977;40:1132–1150. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
