

Functional consequences of neuropeptide and small-molecule co-transmission
- PMID: 28592905
- PMCID: PMC5547741
- DOI: 10.1038/nrn.2017.56
Functional consequences of neuropeptide and small-molecule co-transmission
Abstract
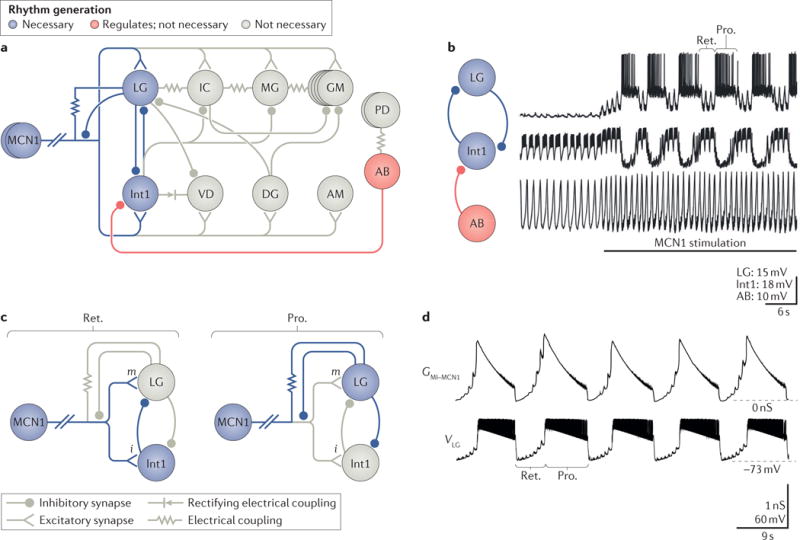
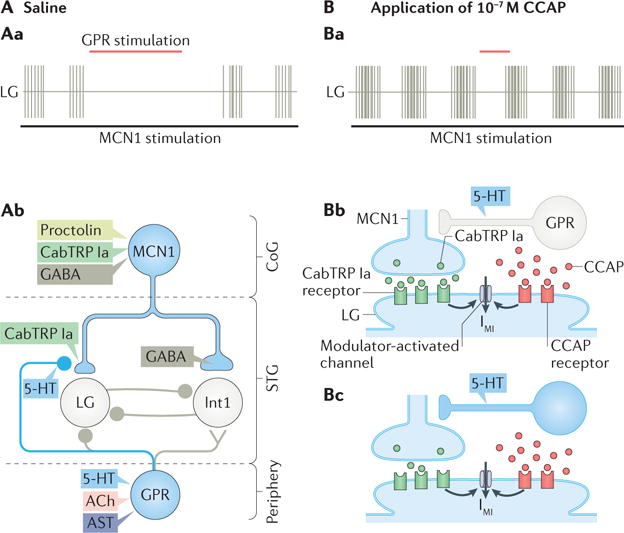
Colocalization of small-molecule and neuropeptide transmitters is common throughout the nervous system of all animals. The resulting co-transmission, which provides conjoint ionotropic ('classical') and metabotropic ('modulatory') actions, includes neuropeptide- specific aspects that are qualitatively different from those that result from metabotropic actions of small-molecule transmitter release. Here, we focus on the flexibility afforded to microcircuits by such co-transmission, using examples from various nervous systems. Insights from such studies indicate that co-transmission mediated even by a single neuron can configure microcircuit activity via an array of contributing mechanisms, operating on multiple timescales, to enhance both behavioural flexibility and robustness.
Conflict of interest statement
The authors declare no competing interests.
Figures
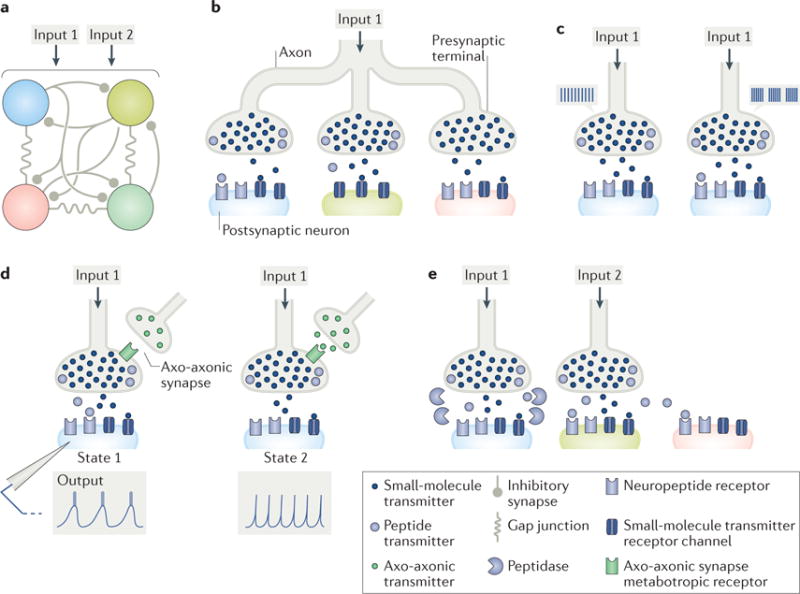
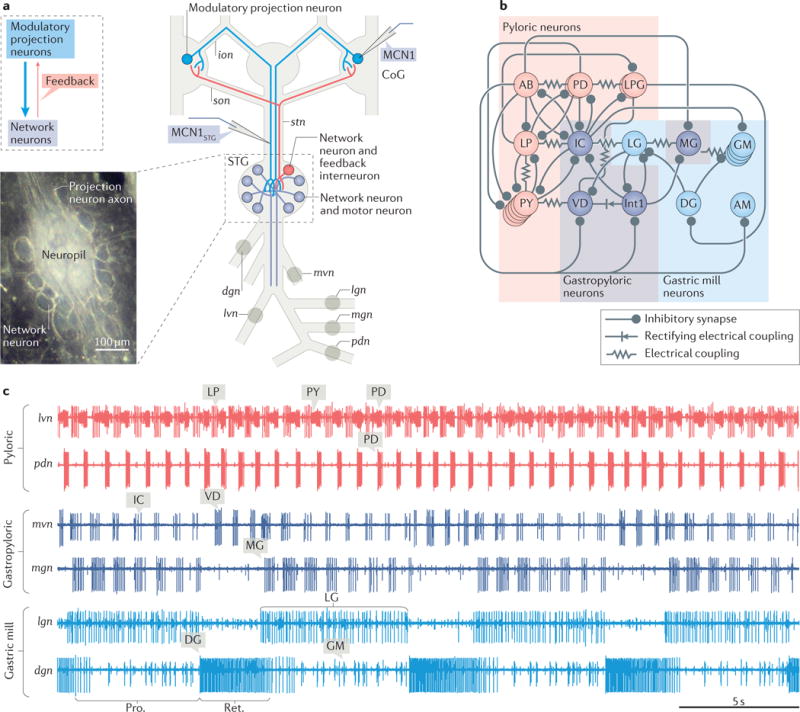
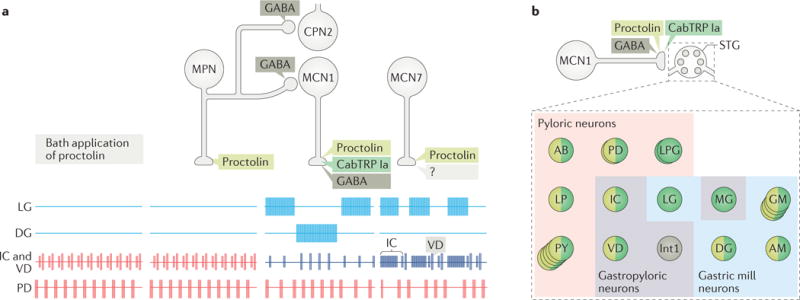
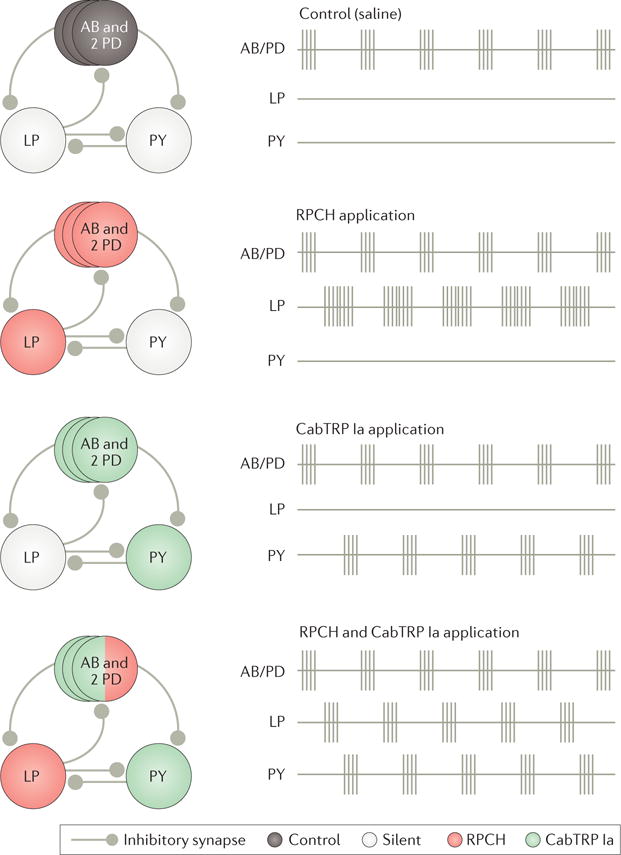
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
