

NS5A inhibitors unmask differences in functional replicase complex half-life between different hepatitis C virus strains
- PMID: 28594932
- PMCID: PMC5464671
- DOI: 10.1371/journal.ppat.1006343
NS5A inhibitors unmask differences in functional replicase complex half-life between different hepatitis C virus strains
Abstract
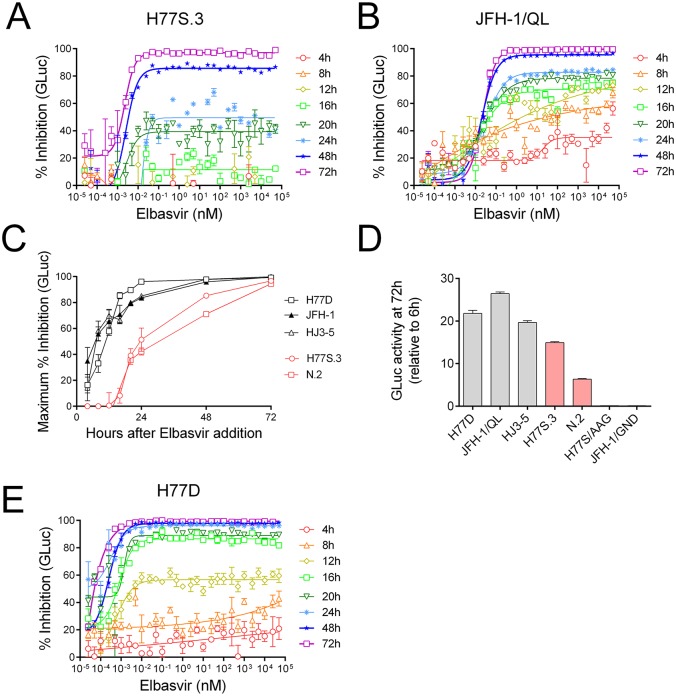
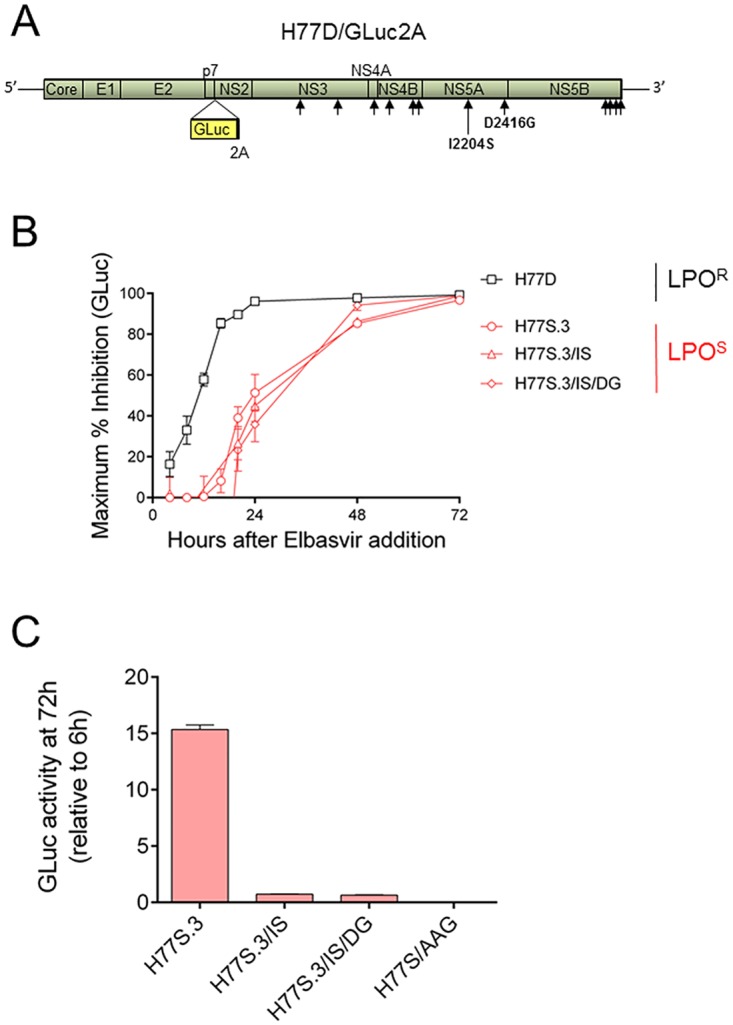
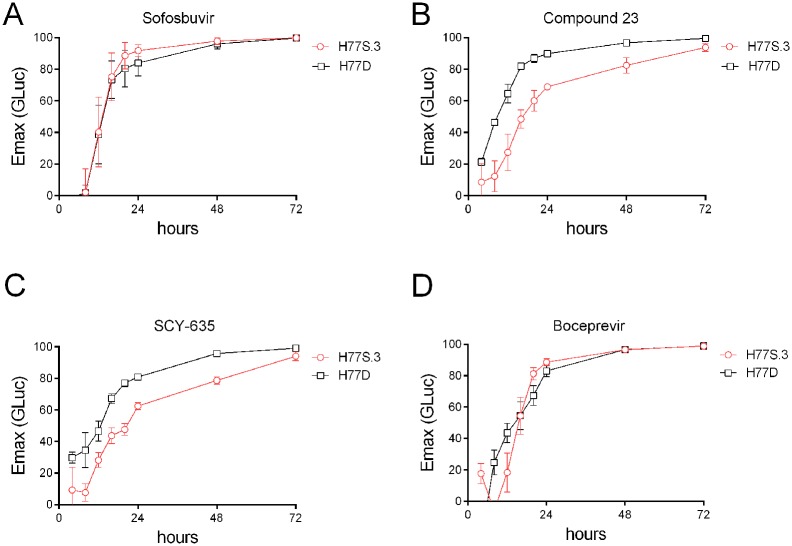
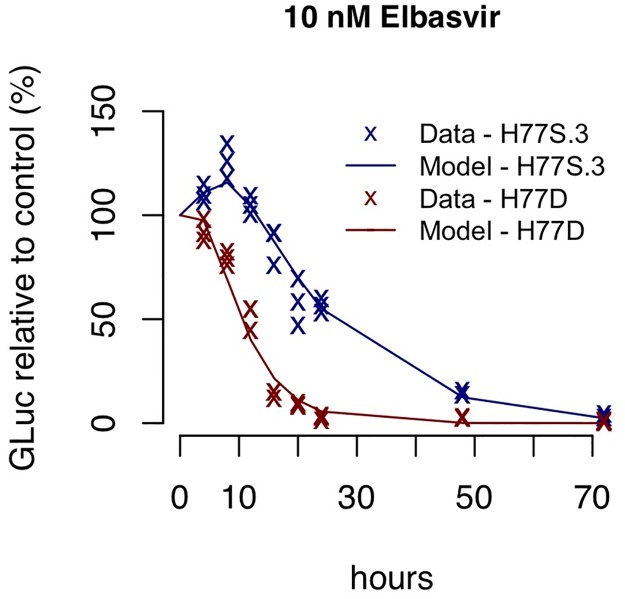
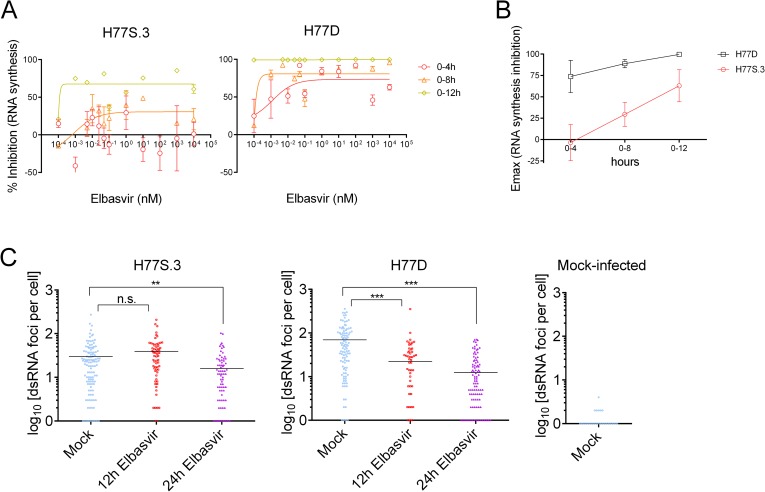
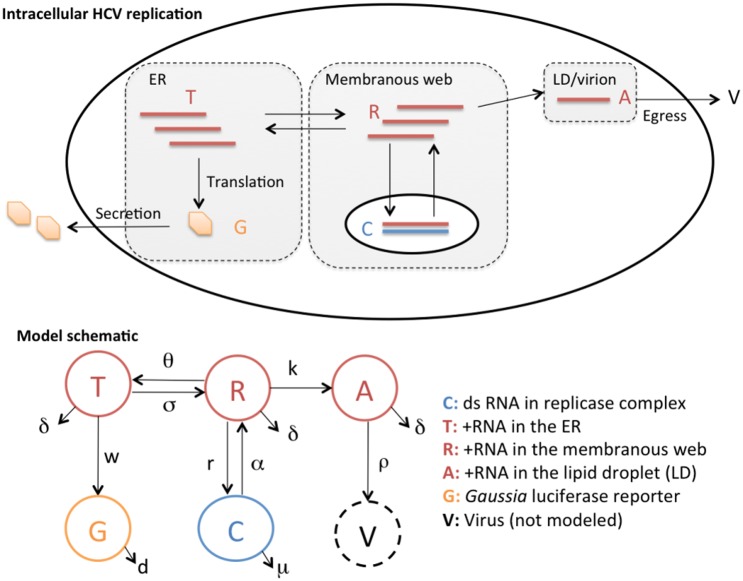
Hepatitis C virus (HCV) RNA is synthesized by the replicase complex (RC), a macromolecular assembly composed of viral non-structural proteins and cellular co-factors. Inhibitors of the HCV NS5A protein block formation of new RCs but do not affect RNA synthesis by pre-formed RCs. Without new RC formation, existing RCs turn over and are eventually lost from the cell. We aimed to use NS5A inhibitors to estimate the half-life of the functional RC of HCV. We compared different cell culture-infectious strains of HCV that may be grouped based on their sensitivity to lipid peroxidation: robustly replicating, lipid peroxidation resistant (LPOR) viruses (e.g. JFH-1 or H77D) and more slowly replicating, lipid peroxidation sensitive (LPOS) viruses (e.g. H77S.3 and N.2). In luciferase assays, LPOS HCV strains declined under NS5A inhibitor therapy with much slower kinetics compared to LPOR HCV strains. This difference in rate of decline was not observed for inhibitors of the NS5B RNA-dependent RNA polymerase suggesting that the difference was not simply a consequence of differences in RNA stability. In further analyses, we compared two isoclonal HCV variants: the LPOS H77S.3 and the LPOR H77D that differ only by 12 amino acids. Differences in rate of decline between H77S.3 and H77D following NS5A inhibitor addition were not due to amino acid sequences in NS5A but rather due to a combination of amino acid differences in the non-structural proteins that make up the HCV RC. Mathematical modeling of intracellular HCV RNA dynamics suggested that differences in RC stability (half-lives of 3.5 and 9.9 hours, for H77D and H77S.3, respectively) are responsible for the different kinetics of antiviral suppression between LPOS and LPOR viruses. In nascent RNA capture assays, the rate of RNA synthesis decline following NS5A inhibitor addition was significantly faster for H77D compared to H77S.3 indicating different half-lives of functional RCs.
Conflict of interest statement
I have read the journal's policy and the authors of this manuscript have the following conflicts of interest: SML has received royalties from AbbVie and grant funding from Merck and Gilead Sciences. DRM has received grant funding from AbbVie and Gilead Sciences. All other authors have no competing interests.
Figures
References
-
- Guedj J, Dahari H, Rong L, Sansone ND, Nettles RE, Cotler SJ, et al. Modeling shows that the NS5A inhibitor daclatasvir has two modes of action and yields a shorter estimate of the hepatitis C virus half-life. Proc Natl Acad Sci U S A. 2013;110(10):3991–6. Epub 2013/02/23. 1203110110 [pii] 10.1073/pnas.1203110110 . - DOI - PMC - PubMed
-
- Berger C, Romero-Brey I, Radujkovic D, Terreux R, Zayas M, Paul D, et al. Daclatasvir-like inhibitors of NS5A block early biogenesis of hepatitis C virus-induced membranous replication factories, independent of RNA replication. Gastroenterology. 2014;147(5):1094–105 e25. 10.1053/j.gastro.2014.07.019 - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
