

ZNF131 suppresses centrosome fragmentation in glioblastoma stem-like cells through regulation of HAUS5
- PMID: 28596487
- PMCID: PMC5564707
- DOI: 10.18632/oncotarget.18153
ZNF131 suppresses centrosome fragmentation in glioblastoma stem-like cells through regulation of HAUS5
Abstract
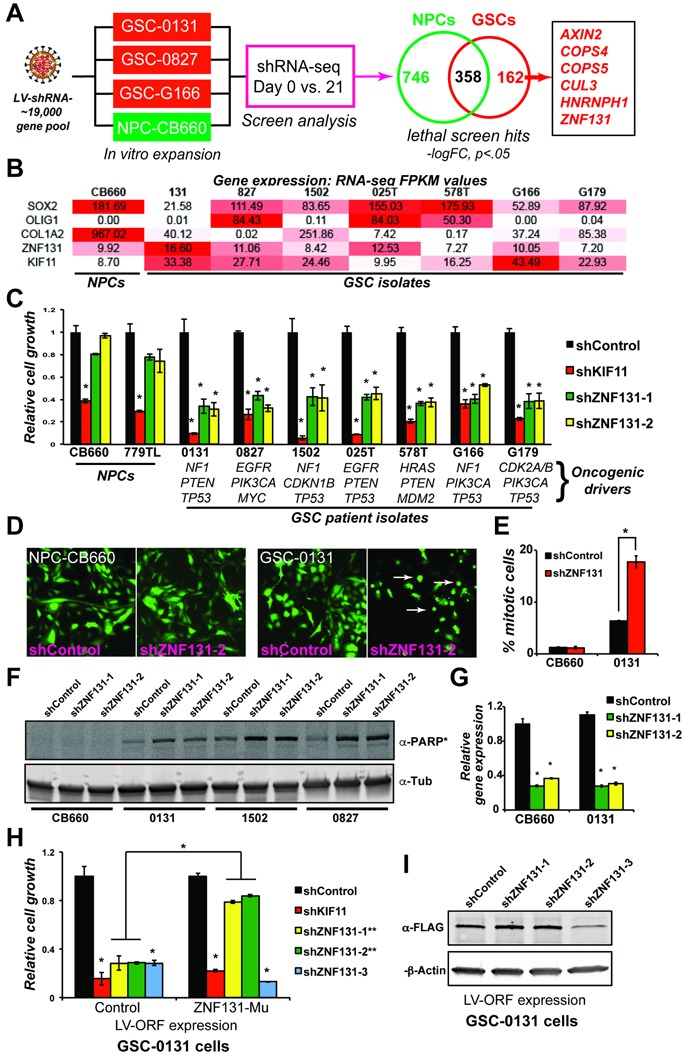
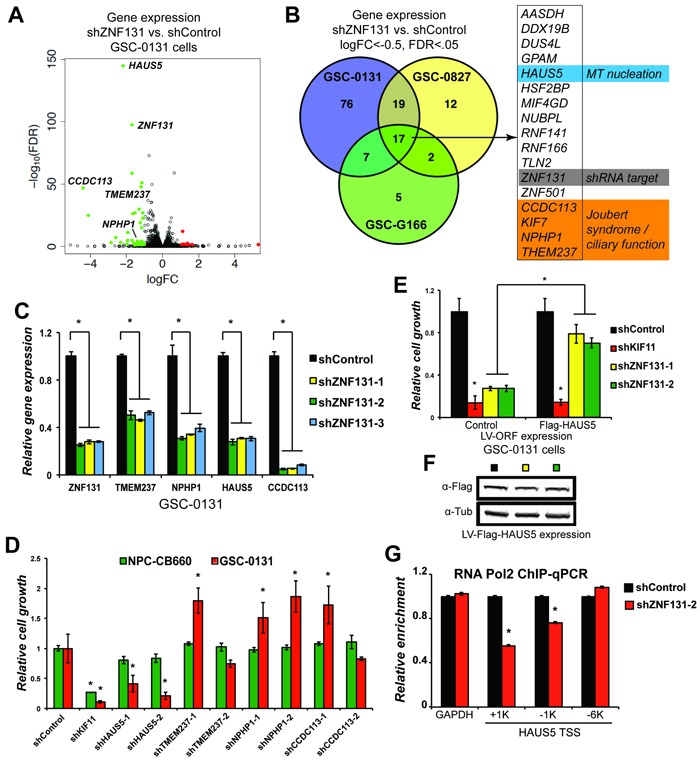
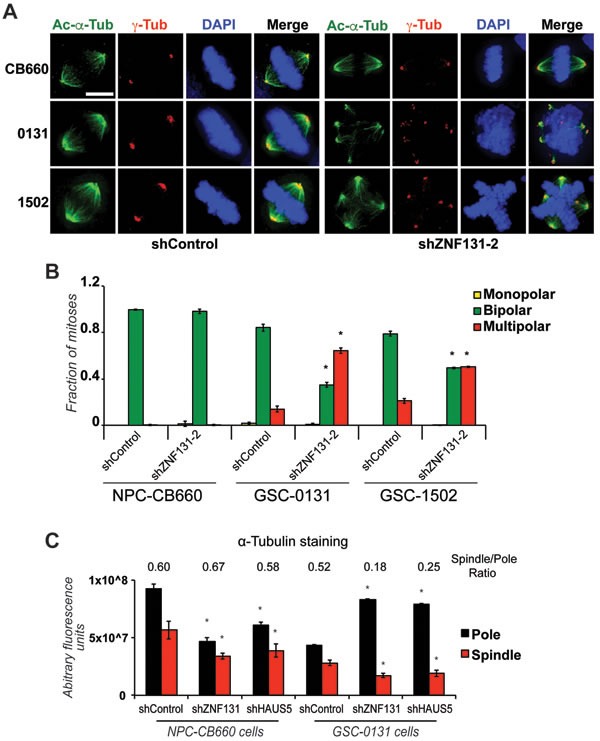
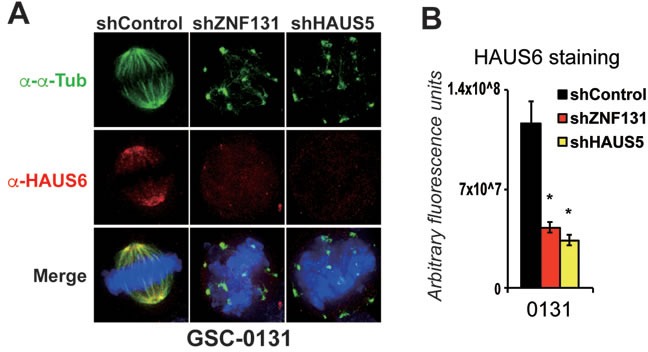
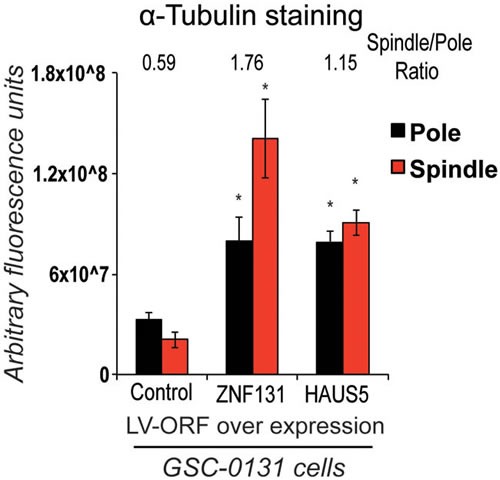
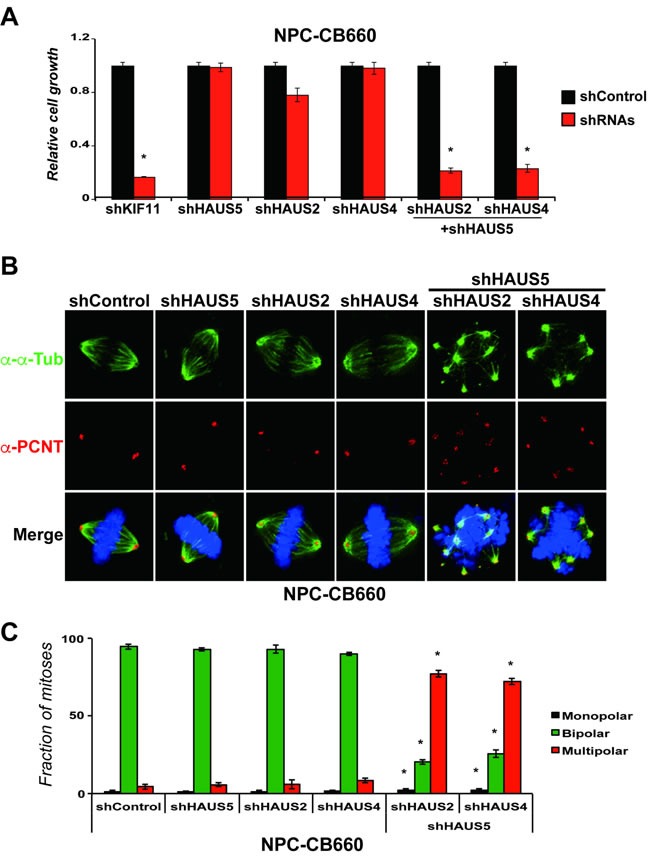
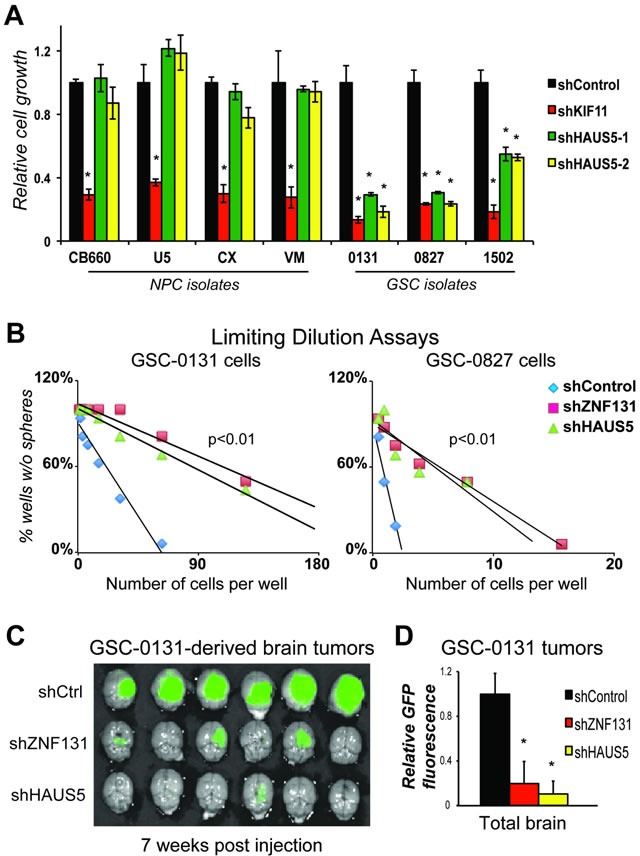
Zinc finger domain genes comprise ~3% of the human genome, yet many of their functions remain unknown. Here we investigated roles for the vertebrate-specific BTB domain zinc finger gene ZNF131 in the context of human brain tumors. We report that ZNF131 is broadly required for Glioblastoma stem-like cell (GSC) viability, but dispensable for neural progenitor cell (NPC) viability. Examination of gene expression changes after ZNF131 knockdown (kd) revealed that ZNF131 activity notably promotes expression of Joubert Syndrome ciliopathy genes, including KIF7, NPHP1, and TMEM237, as well as HAUS5, a component of Augmin/HAUS complex that facilitates microtubule nucleation along the mitotic spindle. Of these genes only kd of HAUS5 displayed GSC-specific viability loss. Critically, HAUS5 ectopic expression was sufficient to suppress viability defects of ZNF131 kd cells. Moreover, ZNF131 and HAUS5 kd phenocopied each other in GSCs, each causing: mitotic arrest, centrosome fragmentation, loss of Augmin/HAUS complex on the mitotic spindle, and loss of GSC self-renewal and tumor formation capacity. In control NPCs, we observed centrosome fragmentation and lethality only when HAUS5 kd was combined with kd of HAUS2 or HAUS4, demonstrating that the complex is essential in NPCs, but that GSCs have heightened requirement. Our results suggest that GSCs differentially rely on ZNF131-dependent expression of HAUS5 as well as the Augmin/HAUS complex activity to maintain the integrity of centrosome function and viability.
Keywords: Augmin/HAUS complex; HAUS5; ZNF131; cancer therapeutics; glioblastoma.
Conflict of interest statement
The authors disclose no potential conflicts of interest.
Figures
References
-
- American Cancer Society American Cancer Society: Cancer Facts and Figures. 2010.
-
- Stupp R, Mason WP, van den Bent MJ, Weller M, Fisher B, Taphoorn MJ, Belanger K, Brandes AA, Marosi C, Bogdahn U, Curschmann J, Janzer RC, Ludwin SK, et al. Radiotherapy plus concomitant and adjuvant temozolomide for glioblastoma. N Engl J Med. 2005;352:987–96. doi: 10.1056/NEJMoa043330. - DOI - PubMed
-
- Singh SK, Clarke ID, Terasaki M, Bonn VE, Hawkins C, Squire J, Dirks PB. Identification of a cancer stem cell in human brain tumors. Cancer Res. 2003;63:5821–8. - PubMed
-
- Singh SK, Hawkins C, Clarke ID, Squire JA, Bayani J, Hide T, Henkelman RM, Cusimano MD, Dirks PB. Identification of human brain tumour initiating cells. Nature. 2004;432:396–401. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
