

Mitochondrial Dysfunction Triggers Synaptic Deficits via Activation of p38 MAP Kinase Signaling in Differentiated Alzheimer's Disease Trans-Mitochondrial Cybrid Cells
- PMID: 28598851
- PMCID: PMC5935489
- DOI: 10.3233/JAD-170283
Mitochondrial Dysfunction Triggers Synaptic Deficits via Activation of p38 MAP Kinase Signaling in Differentiated Alzheimer's Disease Trans-Mitochondrial Cybrid Cells
Abstract
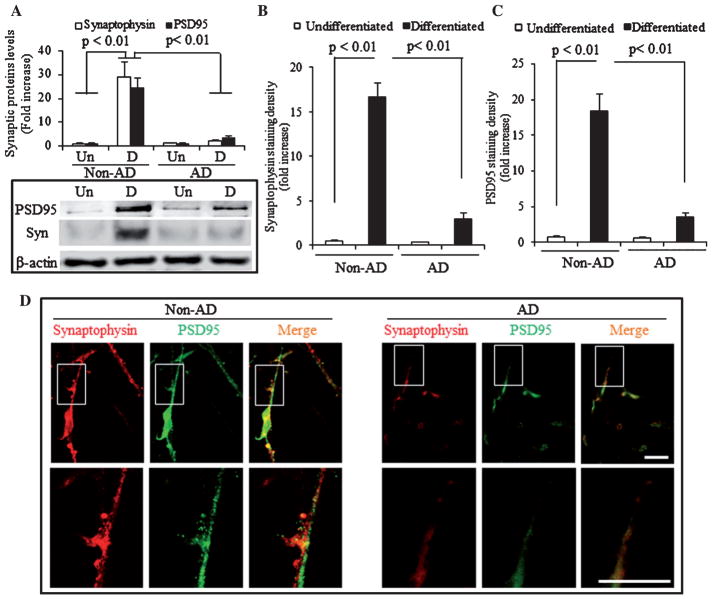
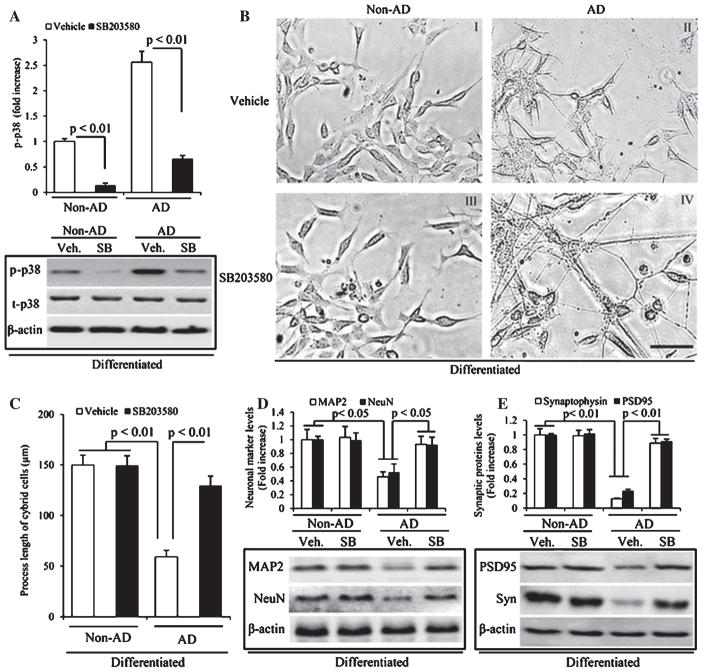
Loss of synapse and synaptic dysfunction contribute importantly to cognitive impairment in Alzheimer's disease (AD). Mitochondrial dysfunction and oxidative stress are early pathological features in AD-affected brain. However, the effect of AD mitochondria on synaptogenesis remains to be determined. Using human trans-mitochondrial "cybrid" (cytoplasmic hybrid) neuronal cells whose mitochondria were transferred from platelets of patients with sporadic AD or age-matched non-AD subjects with relatively normal cognition, we provide the first evidence of mitochondrial dysfunction compromises synaptic development and formation of synapse in AD cybrid cells in response to chemical-induced neuronal differentiation. Compared to non-AD control cybrids, AD cybrid cells showed synaptic loss which was evidenced by a significant reduction in expression of two synaptic marker proteins: synaptophysin (presynaptic marker) and postsynaptic density protein-95, and neuronal proteins (MAP-2 and NeuN) upon neuronal differentiation. In parallel, AD-mediated synaptic deficits correlate to mitochondrial dysfunction and oxidative stress as well as activation of p38 MAP kinase. Notably, inhibition of p38 MAP kinase by pharmacological specific p38 inhibitor significantly increased synaptic density, improved mitochondrial function, and reduced oxidative stress. These results suggest that activation of p38 MAP kinase signaling pathway contributes to AD-mediated impairment in neurogenesis, possibly by inhibiting the neuronal differentiation. Our results provide new insight into the crosstalk of dysfunctional AD mitochondria to synaptic formation and maturation via activation of p38 MAP kinase. Therefore, blockade of p38 MAP kinase signal transduction could be a potential therapeutic strategy for AD by alleviating loss of synapses.
Keywords: Alzheimer’s disease; cybrid cells; mitochondrial dysfunction; synaptic deficits.
Figures




Similar articles
-
PINK1 Activation Attenuates Impaired Neuronal-Like Differentiation and Synaptogenesis and Mitochondrial Dysfunction in Alzheimer's Disease Trans-Mitochondrial Cybrid Cells.J Alzheimers Dis. 2021;81(4):1749-1761. doi: 10.3233/JAD-210095. J Alzheimers Dis. 2021. PMID: 33998543 Free PMC article.
-
Inhibition of ERK-DLP1 signaling and mitochondrial division alleviates mitochondrial dysfunction in Alzheimer's disease cybrid cell.Biochim Biophys Acta. 2014 Feb;1842(2):220-31. doi: 10.1016/j.bbadis.2013.11.009. Epub 2013 Nov 16. Biochim Biophys Acta. 2014. PMID: 24252614 Free PMC article.
-
Antioxidants Rescue Mitochondrial Transport in Differentiated Alzheimer's Disease Trans-Mitochondrial Cybrid Cells.J Alzheimers Dis. 2016 Sep 6;54(2):679-90. doi: 10.3233/JAD-160532. J Alzheimers Dis. 2016. PMID: 27567872 Free PMC article.
-
Mechanism of Oxidative Stress and Synapse Dysfunction in the Pathogenesis of Alzheimer's Disease: Understanding the Therapeutics Strategies.Mol Neurobiol. 2016 Jan;53(1):648-661. doi: 10.1007/s12035-014-9053-6. Epub 2014 Dec 17. Mol Neurobiol. 2016. PMID: 25511446 Free PMC article. Review.
-
Mitochondrial Regulatory Pathways in the Pathogenesis of Alzheimer's Disease.J Alzheimers Dis. 2016 Jul 6;53(4):1257-70. doi: 10.3233/JAD-150967. J Alzheimers Dis. 2016. PMID: 27392851 Review.
Cited by
-
The Role of Mitonuclear Incompatibility in Bipolar Disorder Susceptibility and Resilience Against Environmental Stressors.Front Genet. 2021 Mar 16;12:636294. doi: 10.3389/fgene.2021.636294. eCollection 2021. Front Genet. 2021. PMID: 33815470 Free PMC article. Review.
-
Glucocorticoid-driven mitochondrial damage stimulates Tau pathology.Brain. 2023 Oct 3;146(10):4378-4394. doi: 10.1093/brain/awad127. Brain. 2023. PMID: 37070763 Free PMC article.
-
Chemical Knockdown of Phosphorylated p38 Mitogen-Activated Protein Kinase (MAPK) as a Novel Approach for the Treatment of Alzheimer's Disease.ACS Cent Sci. 2023 Mar 1;9(3):417-426. doi: 10.1021/acscentsci.2c01369. eCollection 2023 Mar 22. ACS Cent Sci. 2023. PMID: 36968534 Free PMC article.
-
Amylin Pharmacology in Alzheimer's Disease Pathogenesis and Treatment.Curr Neuropharmacol. 2022;20(10):1894-1907. doi: 10.2174/1570159X19666211201093147. Curr Neuropharmacol. 2022. PMID: 34852745 Free PMC article. Review.
-
Hippocampal mutant APP and amyloid beta-induced cognitive decline, dendritic spine loss, defective autophagy, mitophagy and mitochondrial abnormalities in a mouse model of Alzheimer's disease.Hum Mol Genet. 2018 Apr 15;27(8):1332-1342. doi: 10.1093/hmg/ddy042. Hum Mol Genet. 2018. PMID: 29408999 Free PMC article.
References
-
- Hardy J, Selkoe DJ. The amyloid hypothesis of Alzheimer’s disease: Progress and problems on the road to therapeutics. Science. 2002;297:353–356. - PubMed
-
- DeKosky ST, Scheff SW. Synapse loss in frontal cortex biopsies in Alzheimer’s disease: Correlation with cognitive severity. Ann Neurol. 1990;27:457–464. - PubMed
-
- Hamos JE, DeGennaro LJ, Drachman DA. Synaptic loss in Alzheimer’s disease and other dementias. Neurology. 1989;39:355–361. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
