

Regulatory Functions of Cellular Energy Sensor SNF1-Related Kinase1 for Leaf Senescence Delay through ETHYLENE- INSENSITIVE3 Repression
- PMID: 28600557
- PMCID: PMC5466610
- DOI: 10.1038/s41598-017-03506-1
Regulatory Functions of Cellular Energy Sensor SNF1-Related Kinase1 for Leaf Senescence Delay through ETHYLENE- INSENSITIVE3 Repression
Abstract
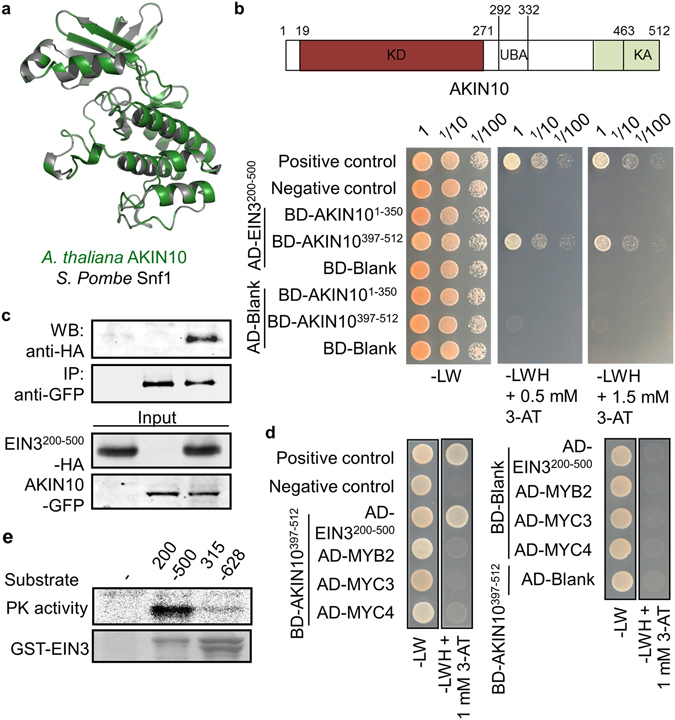
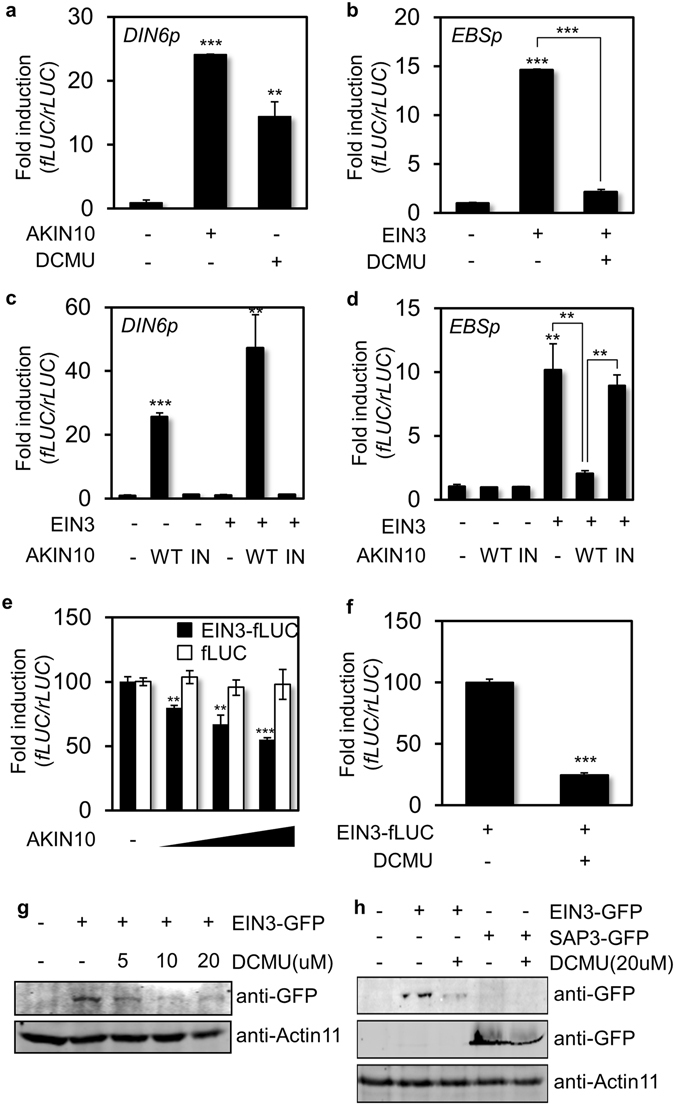
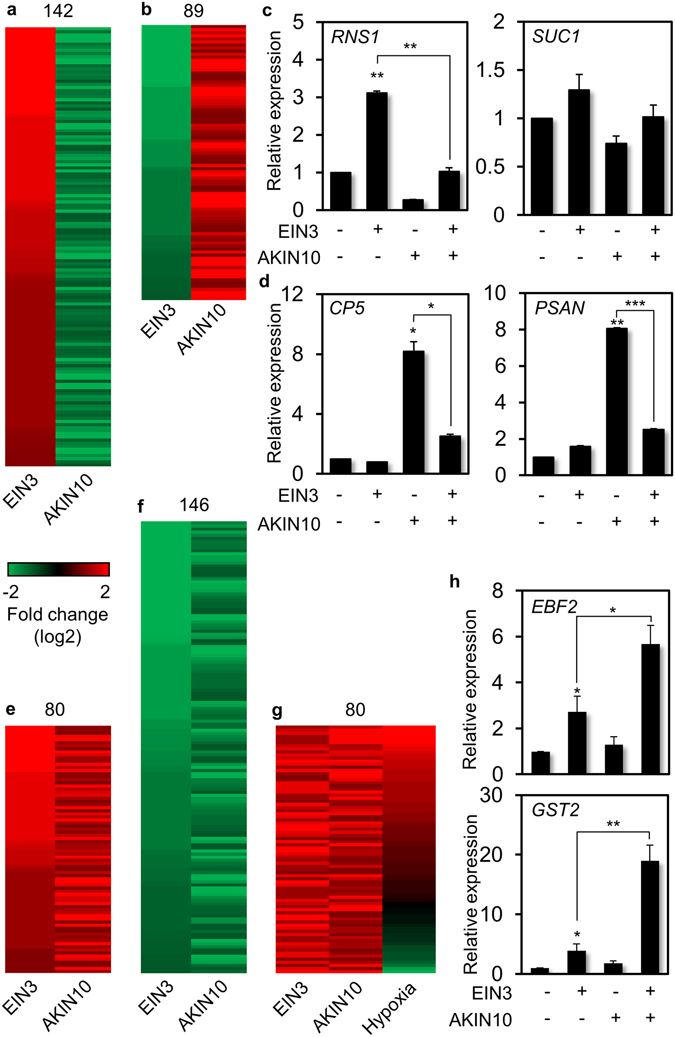
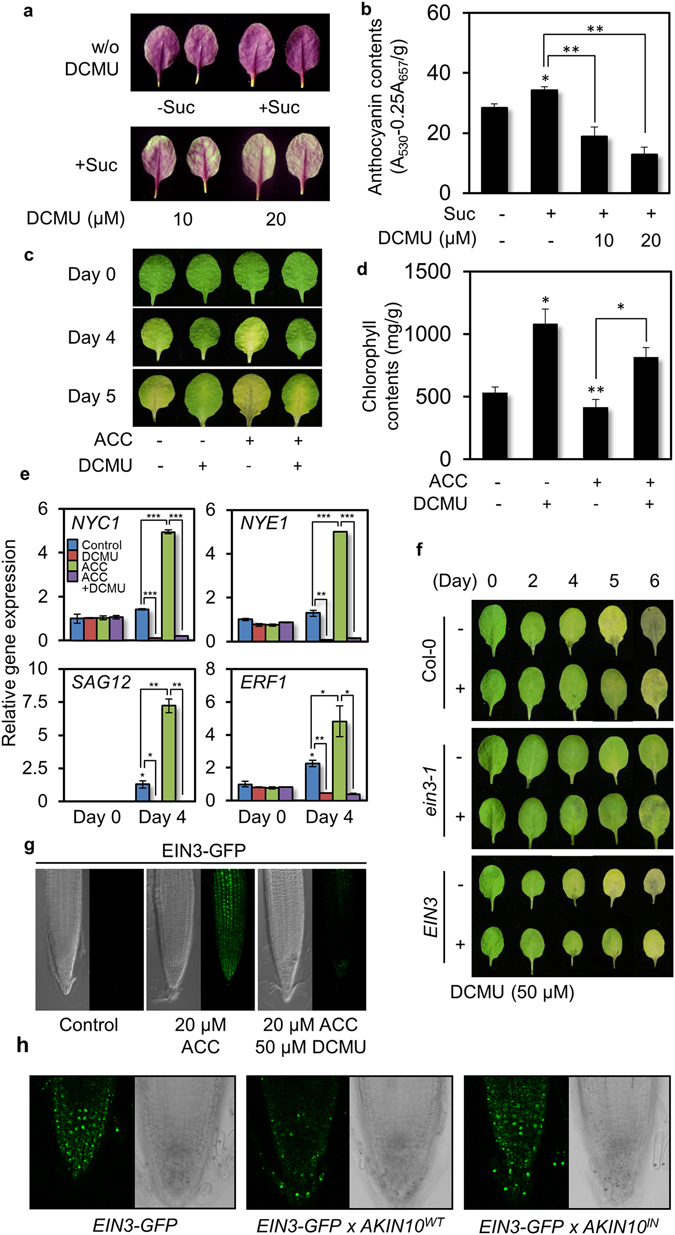
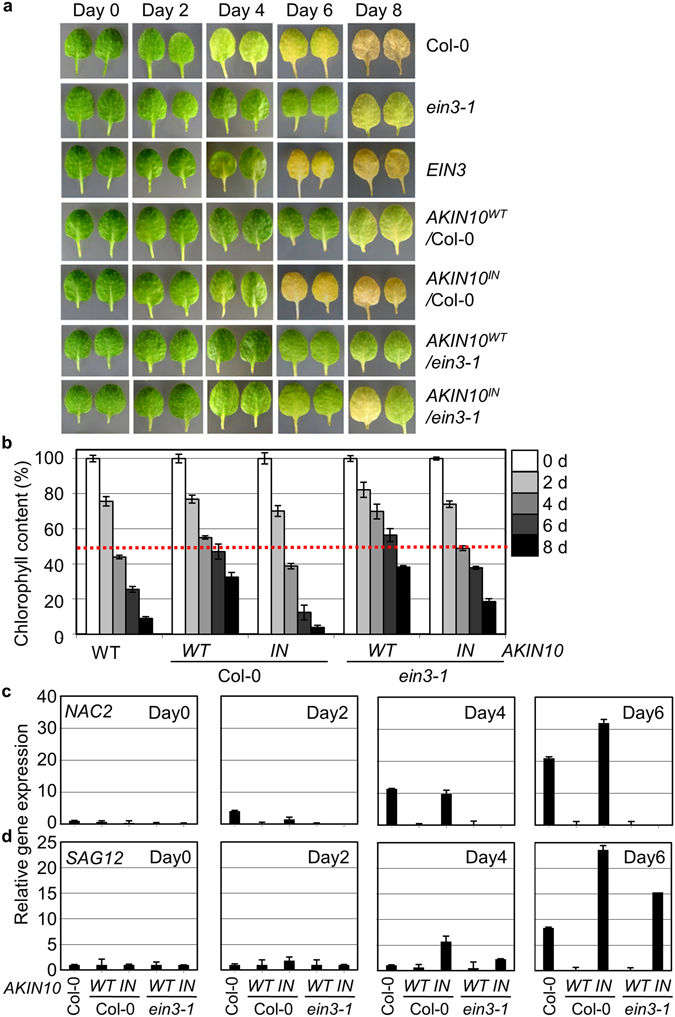
Aging of living organisms is governed by intrinsic developmental programs, of which progression is often under the regulation of their cellular energy status. For example, calorie restriction is known to slow down aging of heterotrophic organisms from yeasts to mammals. In autotrophic plants cellular energy deprivation by perturbation of photosynthesis or sugar metabolism is also shown to induce senescence delay. However, the underlying molecular and biochemical mechanisms remain elusive. Our plant cell-based functional and biochemical assays have demonstrated that SNF1-RELATED KINASE1 (SnRK1) directly interacts, phosphorylates, and destabilizes the key transcription factor ETHYLENE INSENSITIVE3 (EIN3) in senescence-promoting hormone ethylene signaling. Combining chemical manipulation and genetic validation using extended loss-of-function mutants and gain-of-function transgenic lines, we further revealed that a SnRK1 elicitor, 3-(3,4-dichlorophenyl)-1,1-dimethylurea enables to slow down senescence-associated leaf degreening through the regulation of EIN3 in Arabidopsis. Our findings enlighten that an evolutionary conserved cellular energy sensor SnRK1 plays a role in fine-tuning of organ senescence progression to avoid sudden death during the last step of leaf growth and development.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures
Similar articles
-
EIN3 and ORE1 Accelerate Degreening during Ethylene-Mediated Leaf Senescence by Directly Activating Chlorophyll Catabolic Genes in Arabidopsis.PLoS Genet. 2015 Jul 28;11(7):e1005399. doi: 10.1371/journal.pgen.1005399. eCollection 2015 Jul. PLoS Genet. 2015. PMID: 26218222 Free PMC article.
-
Ethylene-insensitive3 is a senescence-associated gene that accelerates age-dependent leaf senescence by directly repressing miR164 transcription in Arabidopsis.Plant Cell. 2013 Sep;25(9):3311-28. doi: 10.1105/tpc.113.113340. Epub 2013 Sep 24. Plant Cell. 2013. PMID: 24064769 Free PMC article.
-
The AtMINPP Gene, Encoding a Multiple Inositol Polyphosphate Phosphatase, Coordinates a Novel Crosstalk between Phytic Acid Metabolism and Ethylene Signal Transduction in Leaf Senescence.Int J Mol Sci. 2024 Aug 17;25(16):8969. doi: 10.3390/ijms25168969. Int J Mol Sci. 2024. PMID: 39201658 Free PMC article.
-
Plant SnRK1 Kinases: Structure, Regulation, and Function.Exp Suppl. 2016;107:403-438. doi: 10.1007/978-3-319-43589-3_17. Exp Suppl. 2016. PMID: 27812990 Review.
-
Shaping plant development through the SnRK1-TOR metabolic regulators.Curr Opin Plant Biol. 2017 Feb;35:152-157. doi: 10.1016/j.pbi.2016.12.004. Epub 2016 Dec 25. Curr Opin Plant Biol. 2017. PMID: 28027512 Review.
Cited by
-
Cytokinin and abiotic stress tolerance -What has been accomplished and the way forward?Front Genet. 2022 Aug 9;13:943025. doi: 10.3389/fgene.2022.943025. eCollection 2022. Front Genet. 2022. Retraction in: Front Genet. 2024 Mar 19;15:1399259. doi: 10.3389/fgene.2024.1399259. PMID: 36017502 Free PMC article. Retracted. Review.
-
Sugar metabolism as input signals and fuel for leaf senescence.Genes Genomics. 2019 Jul;41(7):737-746. doi: 10.1007/s13258-019-00804-y. Epub 2019 Mar 16. Genes Genomics. 2019. PMID: 30879182 Review.
-
Shaping Ethylene Response: The Role of EIN3/EIL1 Transcription Factors.Front Plant Sci. 2019 Aug 26;10:1030. doi: 10.3389/fpls.2019.01030. eCollection 2019. Front Plant Sci. 2019. PMID: 31507622 Free PMC article. Review.
-
Impaired KIN10 function restores developmental defects in the Arabidopsis trehalose 6-phosphate synthase1 (tps1) mutant.New Phytol. 2022 Jul;235(1):220-233. doi: 10.1111/nph.18104. Epub 2022 Apr 8. New Phytol. 2022. PMID: 35306666 Free PMC article.
-
Dynamic Nutrient Signaling Networks in Plants.Annu Rev Cell Dev Biol. 2021 Oct 6;37:341-367. doi: 10.1146/annurev-cellbio-010521-015047. Epub 2021 Aug 5. Annu Rev Cell Dev Biol. 2021. PMID: 34351784 Free PMC article. Review.
References
-
- Weindruch, R. H. & Walford, R. L. The retardation of ageing and disease by dietary restriction. Springfield, IL, Charles C Thomas Publisher (1988).
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
