

Cryo-EM Reveals How Human Cytoplasmic Dynein Is Auto-inhibited and Activated
- PMID: 28602352
- PMCID: PMC5473941
- DOI: 10.1016/j.cell.2017.05.025
Cryo-EM Reveals How Human Cytoplasmic Dynein Is Auto-inhibited and Activated
Abstract
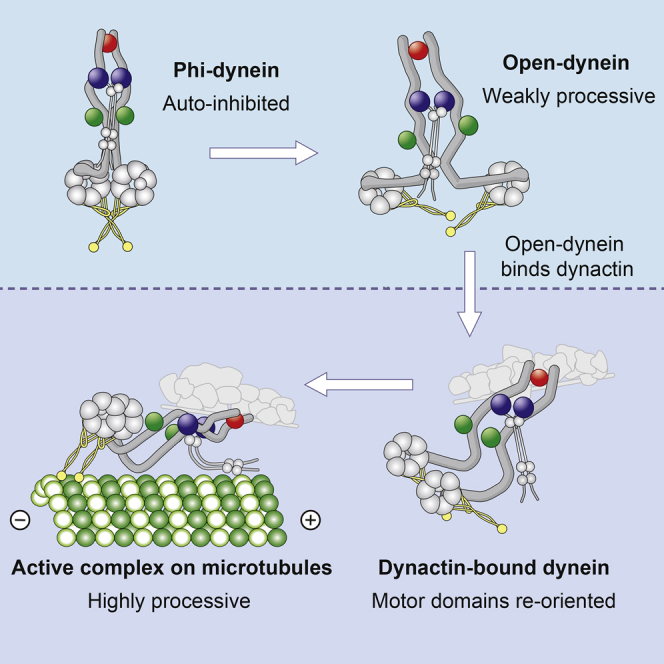
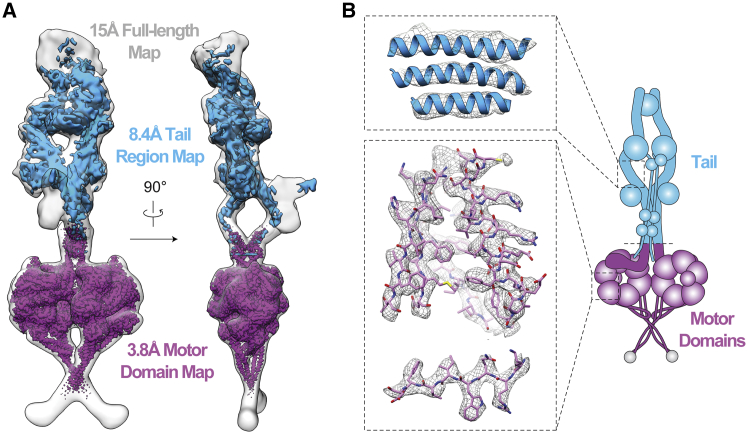
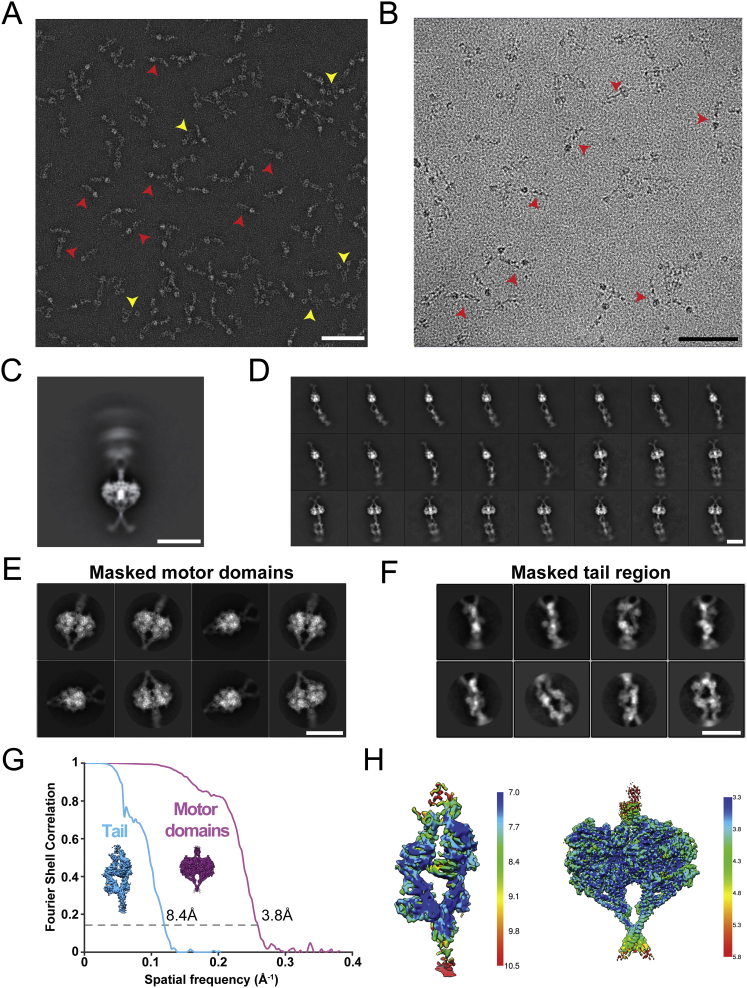
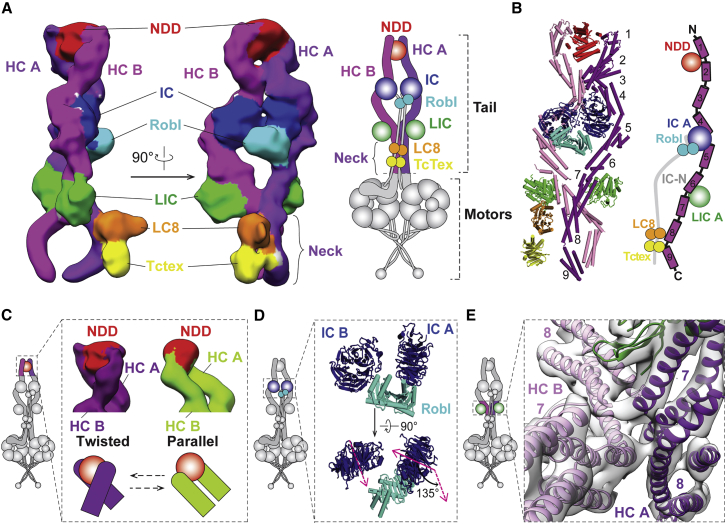
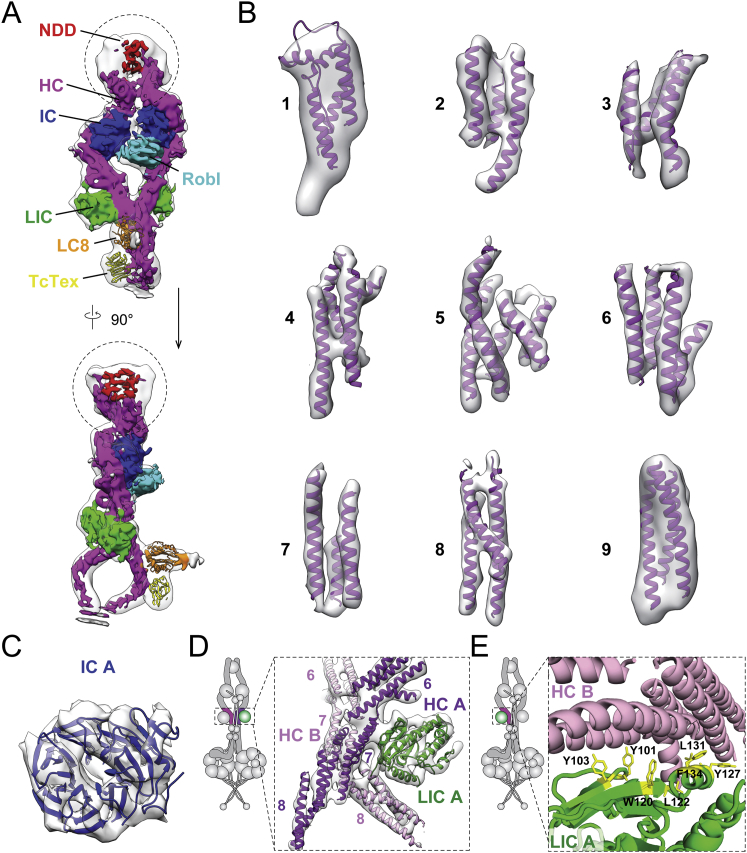
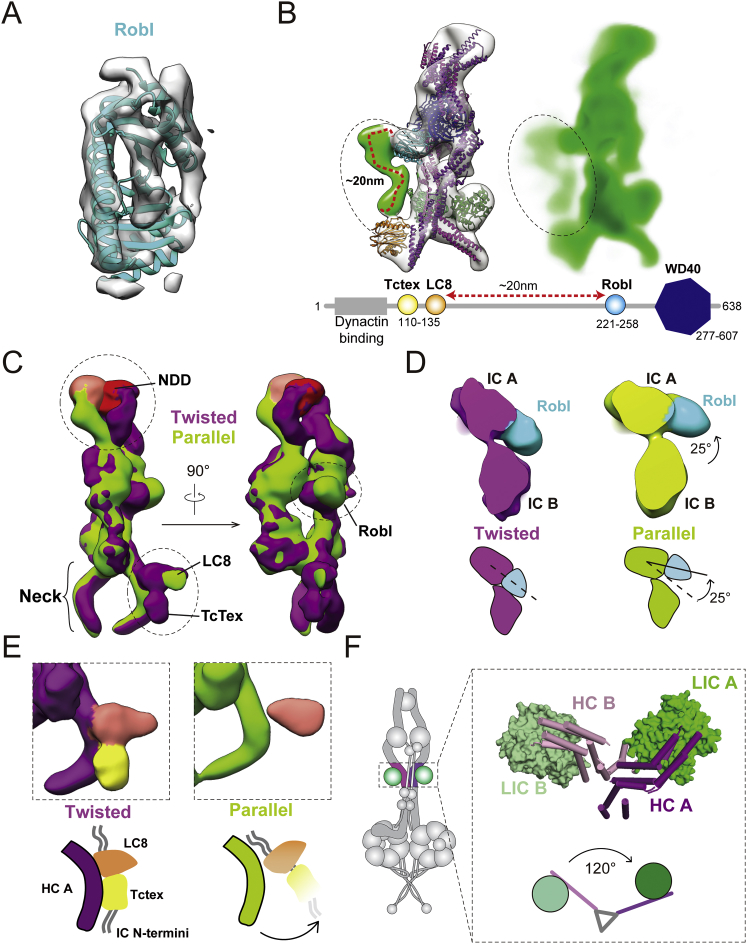
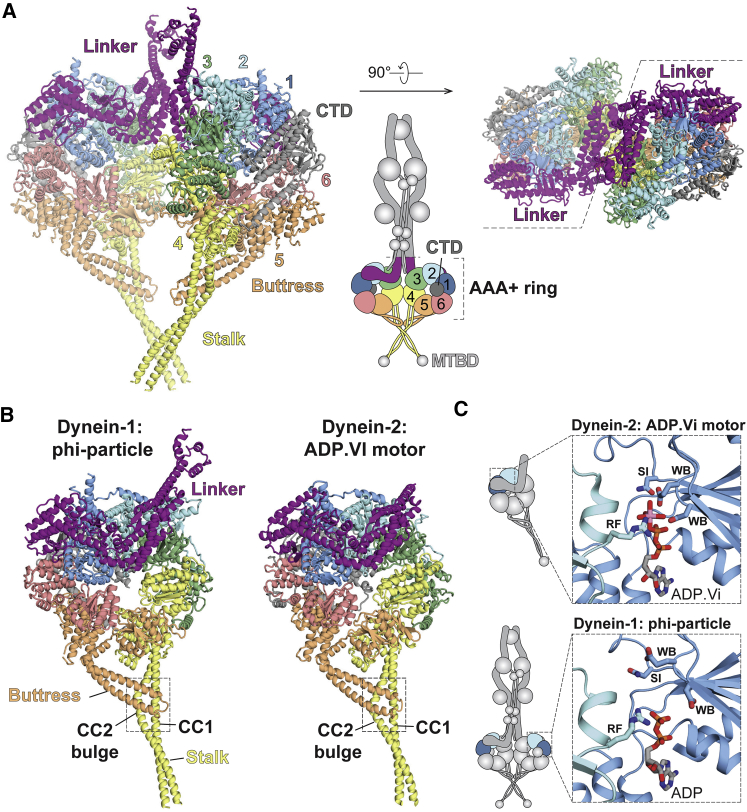
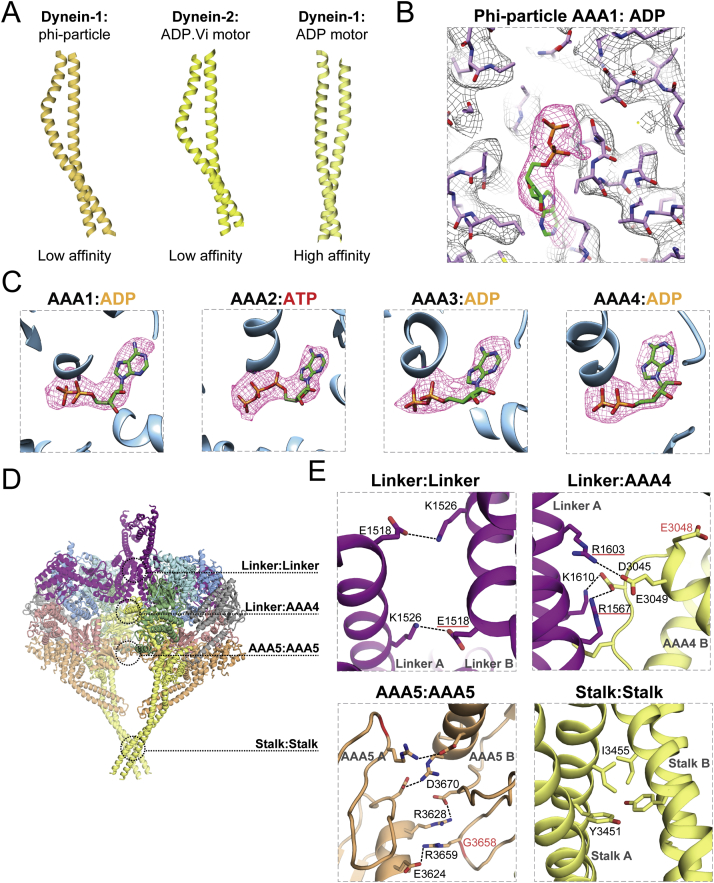
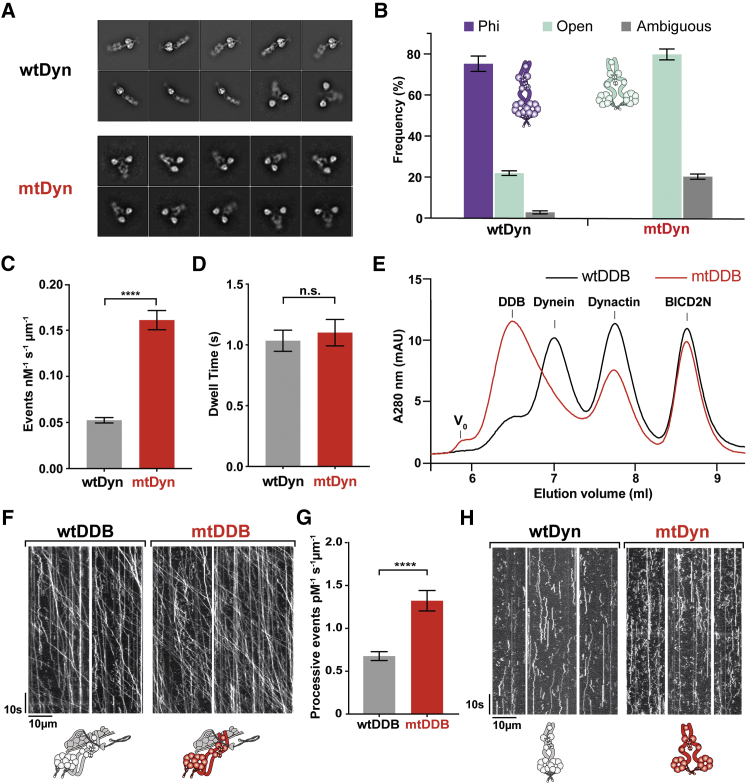
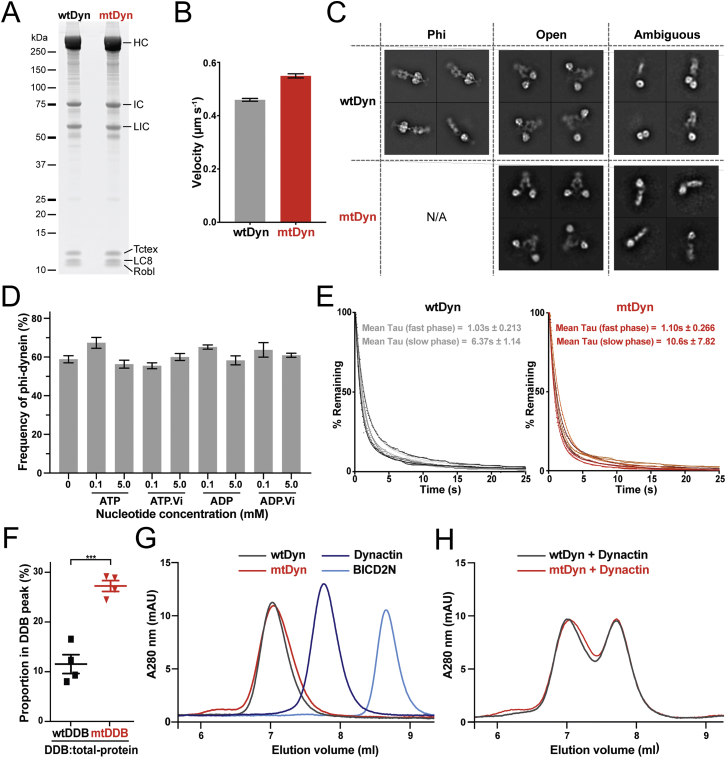
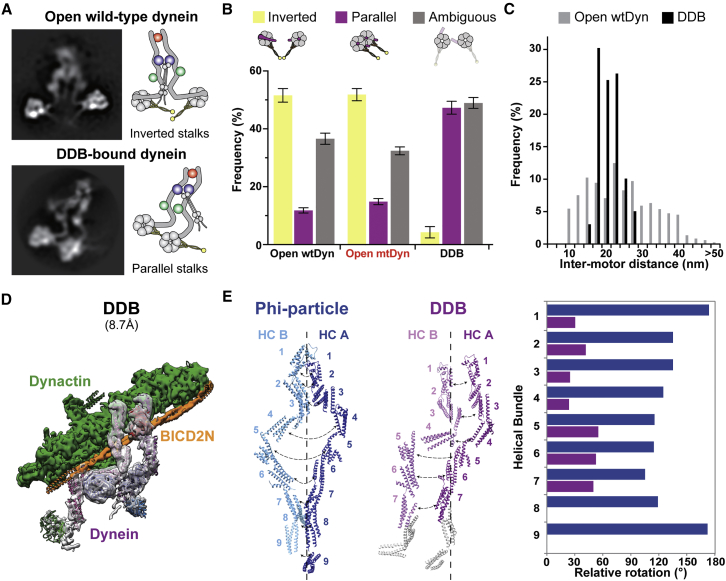
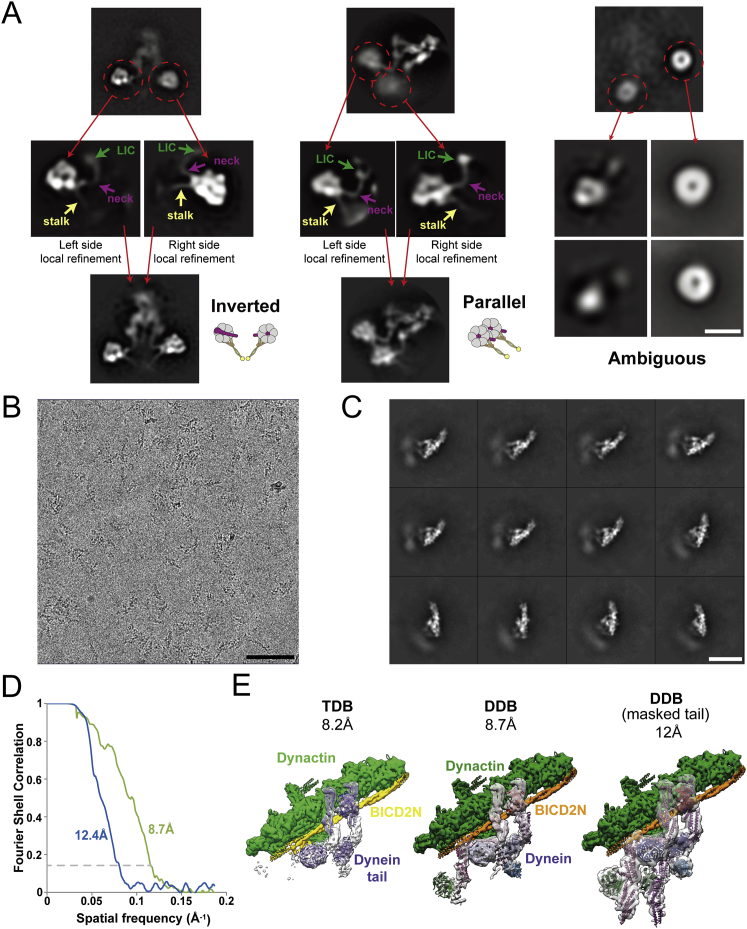
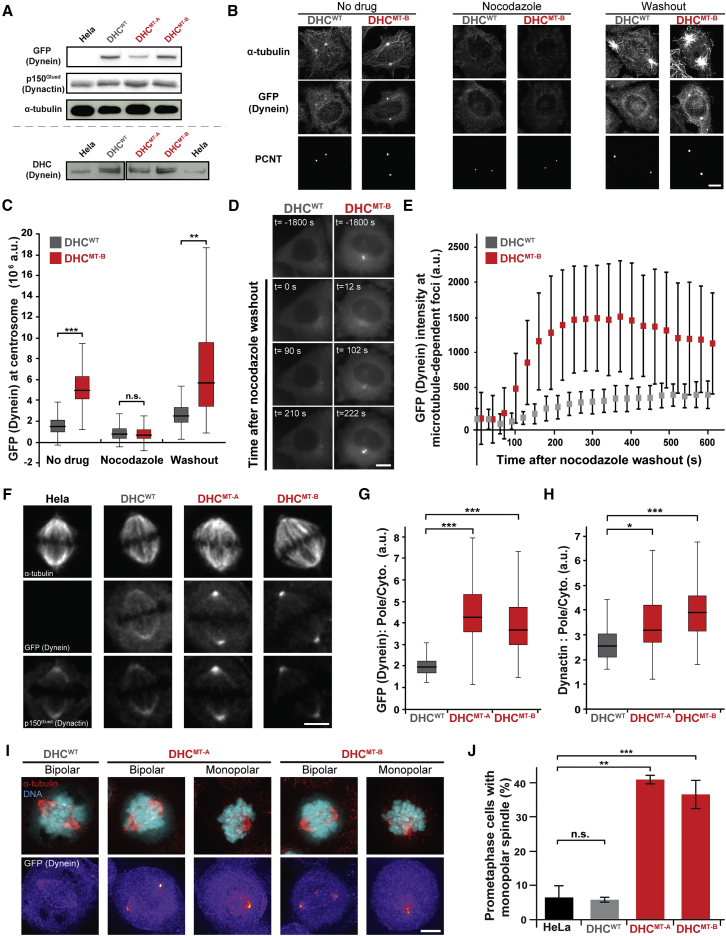
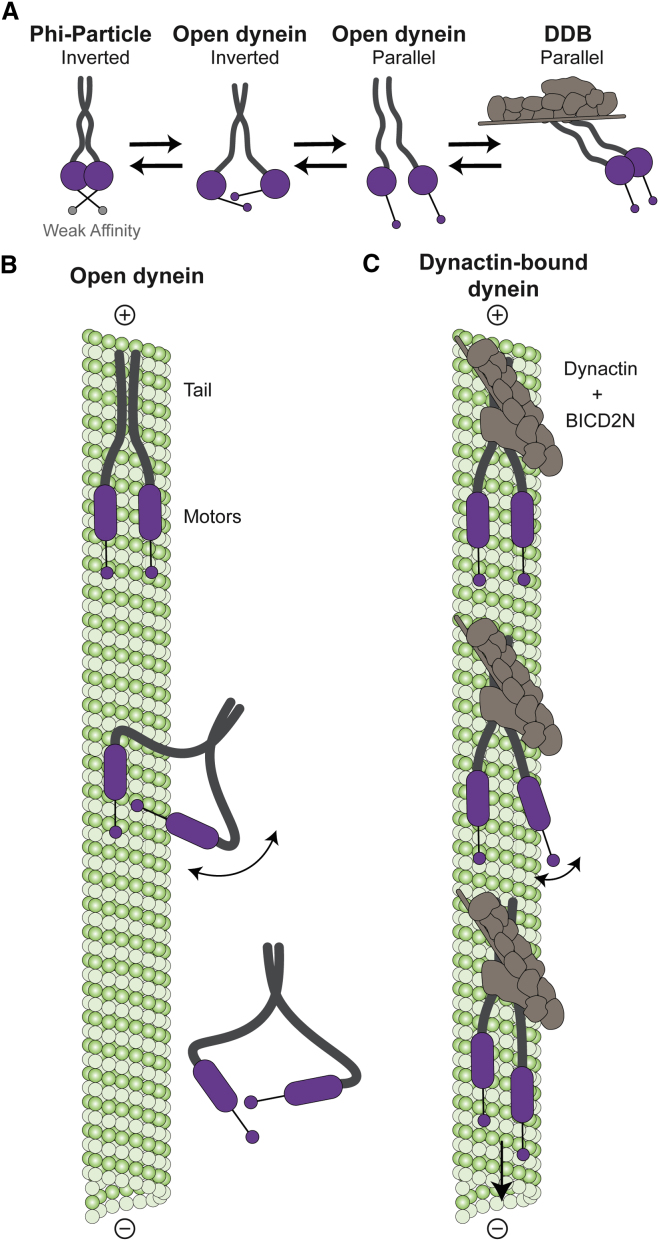
Cytoplasmic dynein-1 binds dynactin and cargo adaptor proteins to form a transport machine capable of long-distance processive movement along microtubules. However, it is unclear why dynein-1 moves poorly on its own or how it is activated by dynactin. Here, we present a cryoelectron microscopy structure of the complete 1.4-megadalton human dynein-1 complex in an inhibited state known as the phi-particle. We reveal the 3D structure of the cargo binding dynein tail and show how self-dimerization of the motor domains locks them in a conformation with low microtubule affinity. Disrupting motor dimerization with structure-based mutagenesis drives dynein-1 into an open form with higher affinity for both microtubules and dynactin. We find the open form is also inhibited for movement and that dynactin relieves this by reorienting the motor domains to interact correctly with microtubules. Our model explains how dynactin binding to the dynein-1 tail directly stimulates its motor activity.
Keywords: activation; auto-inhibition; cryo-EM; dynactin; dynein; microtubule; motor; phi-particle.
Copyright © 2017 MRC Laboratory of Molecular Biology. Published by Elsevier Inc. All rights reserved.
Figures
References
-
- Amos L.A. Brain dynein crossbridges microtubules into bundles. J. Cell Sci. 1989;93:19–28. - PubMed
-
- Bird A.W., Erler A., Fu J., Hériché J.-K., Maresca M., Zhang Y., Hyman A.A., Stewart A.F. High-efficiency counterselection recombineering for site-directed mutagenesis in bacterial artificial chromosomes. Nat. Methods. 2011;9:103–109. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
