

Emerging mechanisms underlying astrogenesis in the developing mammalian brain
- PMID: 28603210
- PMCID: PMC5709539
- DOI: 10.2183/pjab.93.024
Emerging mechanisms underlying astrogenesis in the developing mammalian brain
Abstract
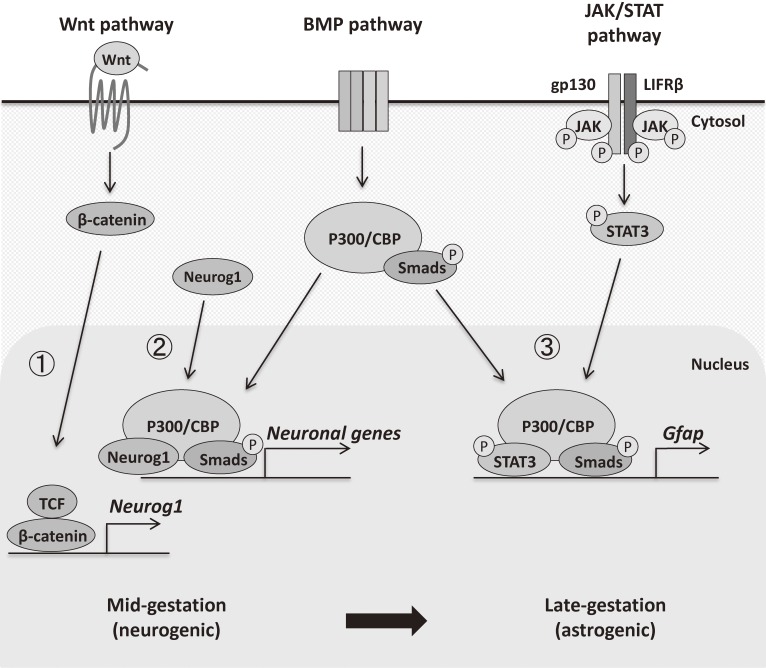
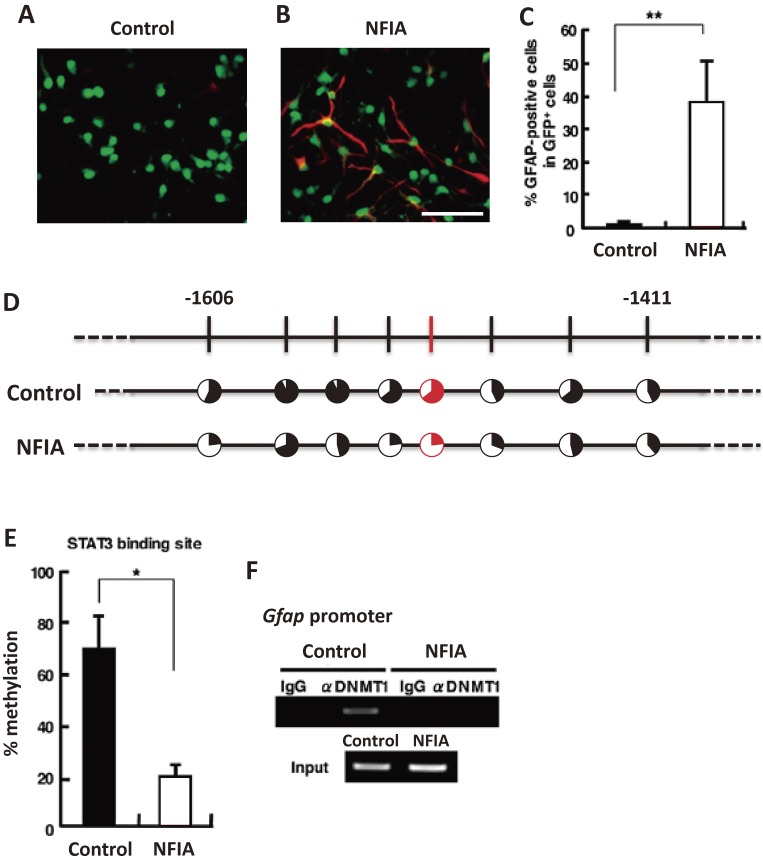
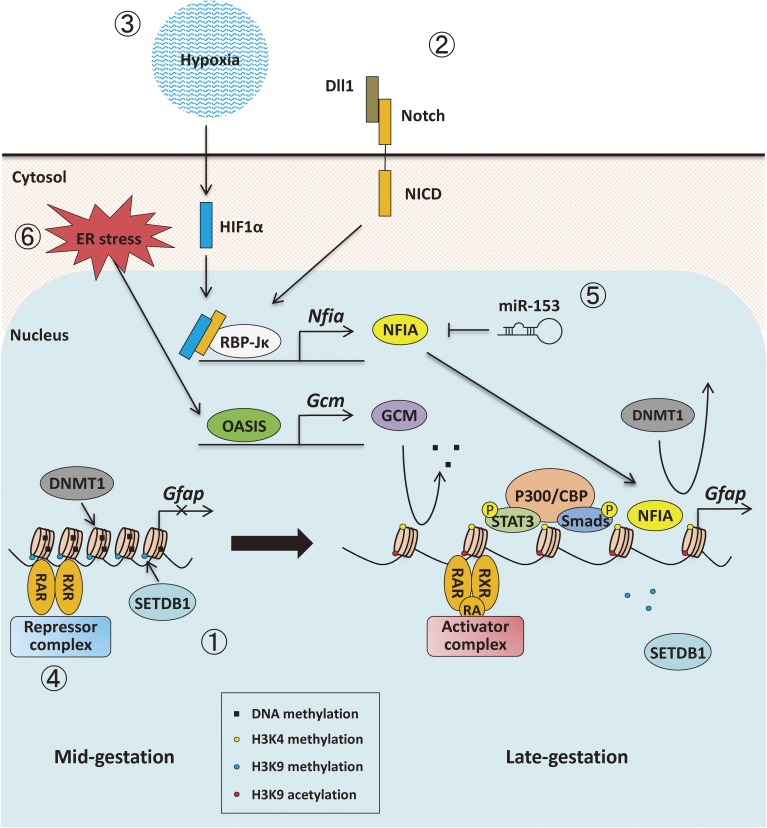
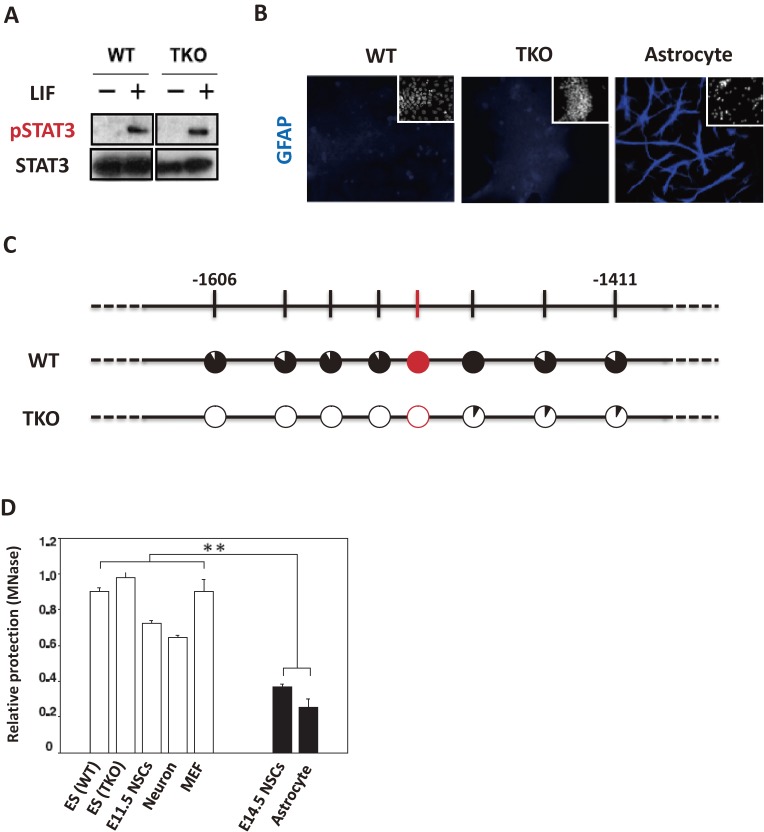
In the developing brain, the three major cell types, i.e., neurons, astrocytes and oligodendrocytes, are generated from common multipotent neural stem cells (NSCs). In particular, astrocytes eventually occupy a great fraction of the brain and play pivotal roles in the brain development and functions. However, NSCs cannot produce the three major cell types simultaneously from the beginning; e.g., it is known that neurogenesis precedes astrogenesis during brain development. How is this fate switching achieved? Many studies have revealed that extracellular cues and intracellular programs are involved in the transition of NSC fate specification. The former include growth factor- and cytokine-signaling, and the latter involve epigenetic machinery, including DNA methylation, histone modifications, and non-coding RNAs. Accumulating evidence has identified a complex array of epigenetic modifications that control the timing of astrocytic differentiation of NSCs. In this review, we introduce recent progress in identifying the molecular mechanisms of astrogenesis underlying the tight regulation of neuronal-astrocytic fate switching of NSCs.
Keywords: astrogenesis; central nerve system (CNS); epigenetics; neural stem cells (NSCs).
Figures
Similar articles
-
Mechanisms of astrocytogenesis in the mammalian brain.Curr Opin Neurobiol. 2013 Dec;23(6):921-7. doi: 10.1016/j.conb.2013.06.002. Epub 2013 Jul 1. Curr Opin Neurobiol. 2013. PMID: 23827784 Review.
-
Epigenetic mechanisms regulating differentiation of neural stem/precursor cells.Epigenomics. 2014;6(6):637-49. doi: 10.2217/epi.14.53. Epigenomics. 2014. PMID: 25531257 Review.
-
A star is born: new insights into the mechanism of astrogenesis.Cell Mol Life Sci. 2014 Feb;71(3):433-47. doi: 10.1007/s00018-013-1435-9. Epub 2013 Aug 2. Cell Mol Life Sci. 2014. PMID: 23907612 Free PMC article. Review.
-
MicroRNAs and epigenetics in adult neurogenesis.Adv Genet. 2014;86:27-44. doi: 10.1016/B978-0-12-800222-3.00002-4. Adv Genet. 2014. PMID: 25172344 Review.
-
Epigenetic regulation in neural stem cell differentiation.Dev Growth Differ. 2010 Aug;52(6):493-504. doi: 10.1111/j.1440-169X.2010.01175.x. Dev Growth Differ. 2010. PMID: 20608952 Review.
Cited by
-
Do astrocytes act as immune cells after pediatric TBI?Neurobiol Dis. 2023 Sep;185:106231. doi: 10.1016/j.nbd.2023.106231. Epub 2023 Jul 17. Neurobiol Dis. 2023. PMID: 37468048 Free PMC article. Review.
-
Epigenetics in blood-brain barrier disruption.Fluids Barriers CNS. 2021 Apr 6;18(1):17. doi: 10.1186/s12987-021-00250-7. Fluids Barriers CNS. 2021. PMID: 33823899 Free PMC article. Review.
-
Gliomas: a reflection of temporal gliogenic principles.Commun Biol. 2024 Feb 6;7(1):156. doi: 10.1038/s42003-024-05833-2. Commun Biol. 2024. PMID: 38321118 Free PMC article. Review.
-
Dual role of FOXG1 in regulating gliogenesis in the developing neocortex via the FGF signalling pathway.Elife. 2025 Mar 14;13:RP101851. doi: 10.7554/eLife.101851. Elife. 2025. PMID: 40085500 Free PMC article.
-
Reactive Gliosis in Neonatal Disorders: Friend or Foe for Neuroregeneration?Cells. 2024 Jan 11;13(2):131. doi: 10.3390/cells13020131. Cells. 2024. PMID: 38247822 Free PMC article. Review.
References
-
- Taupin P., Gage F.H. (2002) Adult neurogenesis and neural stem cells of the central nervous system in mammals. J. Neurosci. Res. 69, 745–749. - PubMed
-
- Azevedo F.A., Carvalho L.R., Grinberg L.T., Farfel J.M., Ferretti R.E., Leite R.E., Jacob Filho W., Lent R., Herculano-Houzel S. (2009) Equal numbers of neuronal and nonneuronal cells make the human brain an isometrically scaled-up primate brain. J. Comp. Neurol. 513, 532–541. - PubMed
-
- Araque A., Parpura V., Sanzgiri R.P., Haydon P.G. (1999) Tripartite synapses: glia, the unacknowledged partner. Trends Neurosci. 22, 208–215. - PubMed
-
- Nedergaard M., Ransom B., Goldman S.A. (2003) New roles for astrocytes: redefining the functional architecture of the brain. Trends Neurosci. 26, 523–530. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources