

The nematode homologue of Mediator complex subunit 28, F28F8.5, is a critical regulator of C. elegans development
- PMID: 28603670
- PMCID: PMC5464003
- DOI: 10.7717/peerj.3390
The nematode homologue of Mediator complex subunit 28, F28F8.5, is a critical regulator of C. elegans development
Abstract
The evolutionarily conserved Mediator complex is a critical player in regulating transcription. Comprised of approximately two dozen proteins, the Mediator integrates diverse regulatory signals through direct protein-protein interactions that, in turn, modulate the influence of Mediator on RNA Polymerase II activity. One Mediator subunit, MED28, is known to interact with cytoplasmic structural proteins, providing a potential direct link between cytoplasmic dynamics and the control of gene transcription. Although identified in many animals and plants, MED28 is not present in yeast; no bona fide MED28 has been described previously in Caenorhabditis elegans. Here, we identify bioinformatically F28F8.5, an uncharacterized predicted protein, as the nematode homologue of MED28. As in other Metazoa, F28F8.5 has dual nuclear and cytoplasmic localization and plays critical roles in the regulation of development. F28F8.5 is a vital gene and its null mutants have severely malformed gonads and do not reproduce. F28F8.5 interacts on the protein level with the Mediator subunits MDT-6 and MDT-30. Our results indicate that F28F8.5 is an orthologue of MED28 and suggest that the potential to link cytoplasmic and nuclear events is conserved between MED28 vertebrate and nematode orthologues.
Keywords: Caenorhabditis elegans; Development; Evolution; Gene expression regulation; MED28; Mediator complex.
Conflict of interest statement
Marta Kostrouchová is an Academic Editor for PeerJ. No other competing interests declared.
Figures


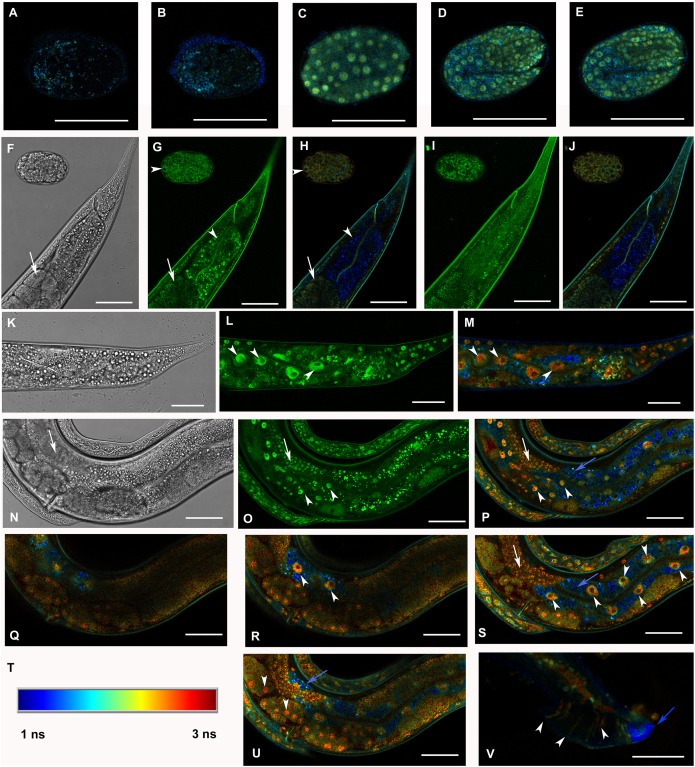




Similar articles
-
Valproic Acid Decreases the Nuclear Localization of MDT-28, the Nematode Orthologue of MED28.Folia Biol (Praha). 2018;64(1):1-9. doi: 10.14712/fb2018064010001. Folia Biol (Praha). 2018. PMID: 29871732
-
Proteomic Interactome of C. elegans Mediator Complex Subunit 28 (MDT-28) Reveals Predominant Association with a Restricted Set of Core Mediator Subunits and an Affinity to Additional Structural and Enzymatic Proteins.Folia Biol (Praha). 2019;65(5-6):203-211. doi: 10.14712/fb2019065050203. Folia Biol (Praha). 2019. PMID: 32362303
-
Mediator subunit MED28 (Magicin) is a repressor of smooth muscle cell differentiation.J Biol Chem. 2007 Nov 2;282(44):32152-7. doi: 10.1074/jbc.M706592200. Epub 2007 Sep 11. J Biol Chem. 2007. PMID: 17848560
-
The Mediator complex of Caenorhabditis elegans: insights into the developmental and physiological roles of a conserved transcriptional coregulator.Nucleic Acids Res. 2015 Feb 27;43(4):2442-53. doi: 10.1093/nar/gkv037. Epub 2015 Jan 29. Nucleic Acids Res. 2015. PMID: 25634893 Free PMC article. Review.
-
Nuclear receptors in nematode development: Natural experiments made by a phylum.Biochim Biophys Acta. 2015 Feb;1849(2):224-37. doi: 10.1016/j.bbagrm.2014.06.016. Epub 2014 Jun 28. Biochim Biophys Acta. 2015. PMID: 24984201 Review.
References
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
