

DNMT and HDAC inhibitors induce cryptic transcription start sites encoded in long terminal repeats
- PMID: 28604729
- PMCID: PMC6005702
- DOI: 10.1038/ng.3889
DNMT and HDAC inhibitors induce cryptic transcription start sites encoded in long terminal repeats
Erratum in
-
Erratum: DNMT and HDAC inhibitors induce cryptic transcription start sites encoded in long terminal repeats.Nat Genet. 2017 Oct 27;49(11):1661. doi: 10.1038/ng1117-1661c. Nat Genet. 2017. PMID: 29074949
Abstract
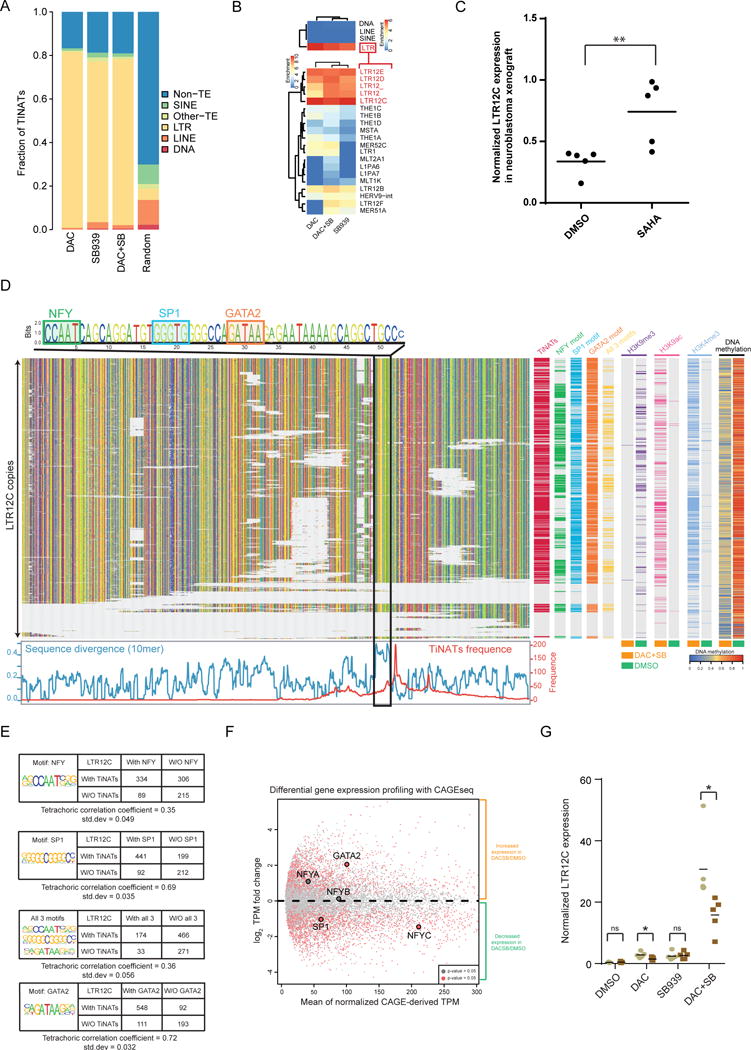
Several mechanisms of action have been proposed for DNA methyltransferase and histone deacetylase inhibitors (DNMTi and HDACi), primarily based on candidate-gene approaches. However, less is known about their genome-wide transcriptional and epigenomic consequences. By mapping global transcription start site (TSS) and chromatin dynamics, we observed the cryptic transcription of thousands of treatment-induced non-annotated TSSs (TINATs) following DNMTi and HDACi treatment. The resulting transcripts frequently splice into protein-coding exons and encode truncated or chimeric ORFs translated into products with predicted abnormal or immunogenic functions. TINAT transcription after DNMTi treatment coincided with DNA hypomethylation and gain of classical promoter histone marks, while HDACi specifically induced a subset of TINATs in association with H2AK9ac, H3K14ac, and H3K23ac. Despite this mechanistic difference, both inhibitors convergently induced transcription from identical sites, as we found TINATs to be encoded in solitary long terminal repeats of the ERV9/LTR12 family, which are epigenetically repressed in virtually all normal cells.
Conflict of interest statement
The authors declare no competing financial interests.
Figures
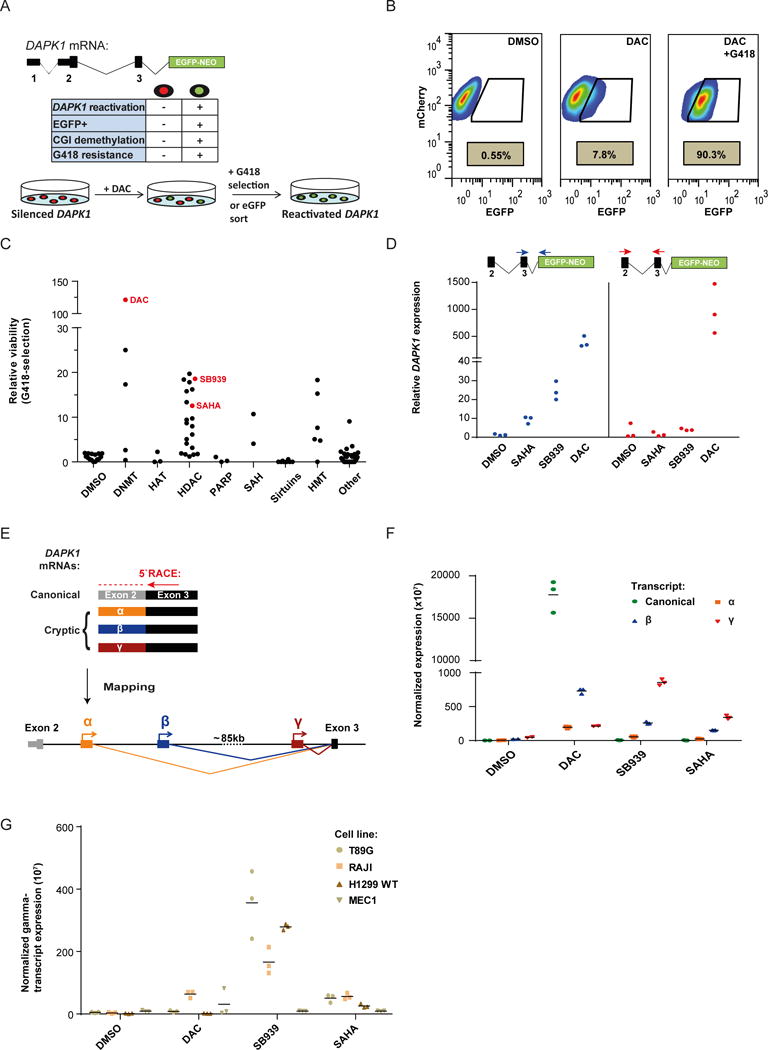
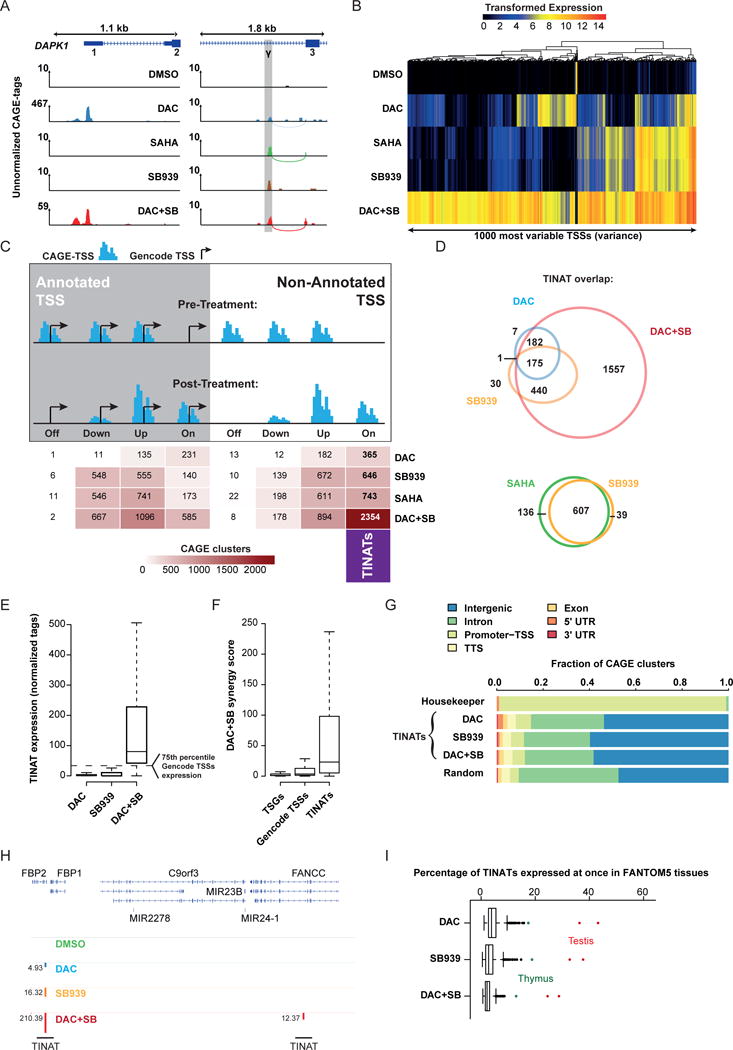
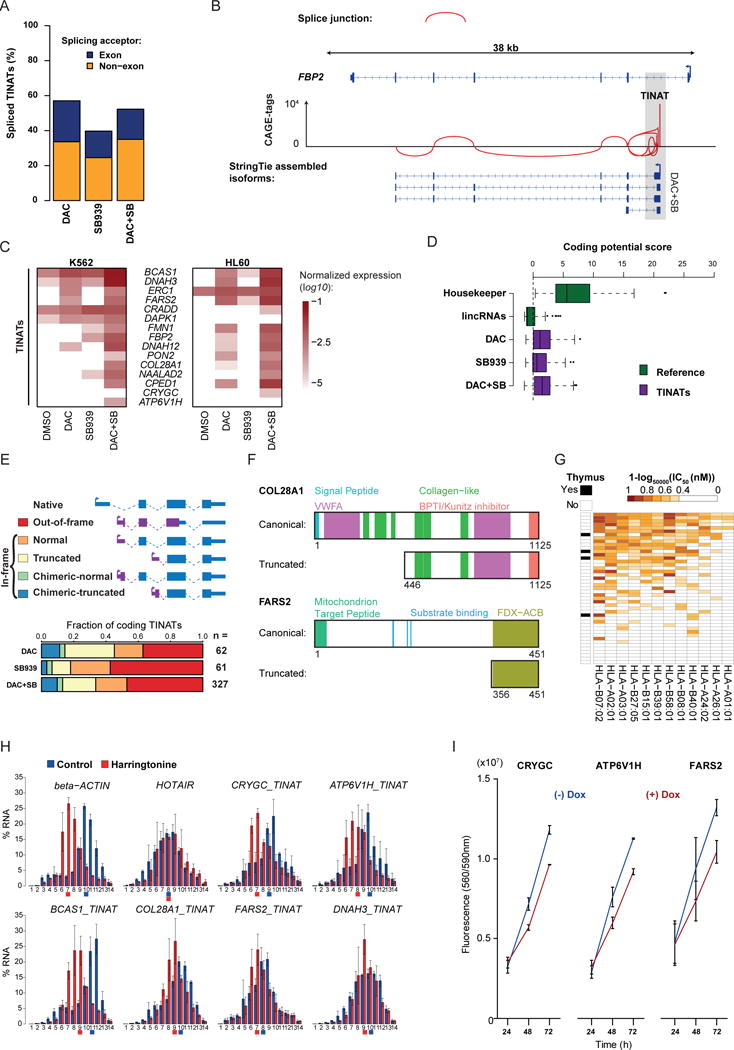
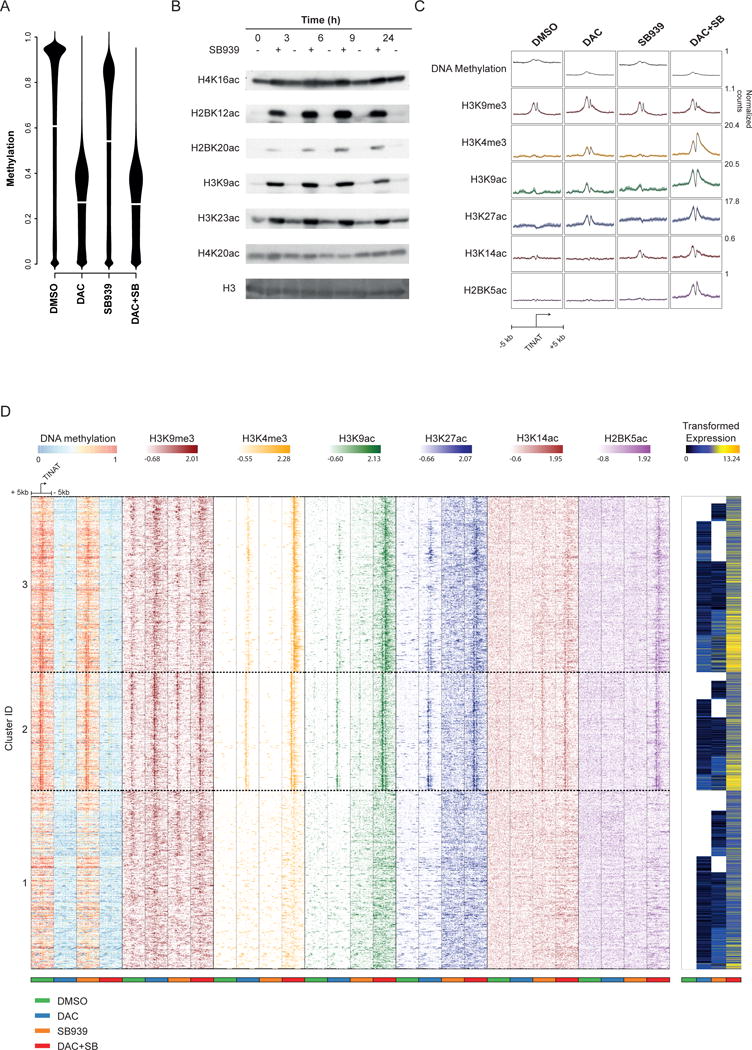
Comment in
-
Cancer epigenetics: Therapy-induced transcription is cryptically widespread.Nat Rev Genet. 2017 Aug;18(8):455. doi: 10.1038/nrg.2017.53. Epub 2017 Jun 26. Nat Rev Genet. 2017. PMID: 28649134 No abstract available.
-
Cancer biology: Therapy-induced transcription is cryptically widespread.Nat Rev Mol Cell Biol. 2017 Aug;18(8):469. doi: 10.1038/nrm.2017.64. Epub 2017 Jun 28. Nat Rev Mol Cell Biol. 2017. PMID: 28655907 No abstract available.
-
Epigenetics: Therapy-induced transcription is cryptically widespread.Nat Rev Cancer. 2017 Aug;17(8):456. doi: 10.1038/nrc.2017.57. Epub 2017 Jul 14. Nat Rev Cancer. 2017. PMID: 28706265 No abstract available.
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
