

The deubiquitinating enzymes USP4 and USP17 target hyaluronan synthase 2 and differentially affect its function
- PMID: 28604766
- PMCID: PMC5519194
- DOI: 10.1038/oncsis.2017.45
The deubiquitinating enzymes USP4 and USP17 target hyaluronan synthase 2 and differentially affect its function
Abstract
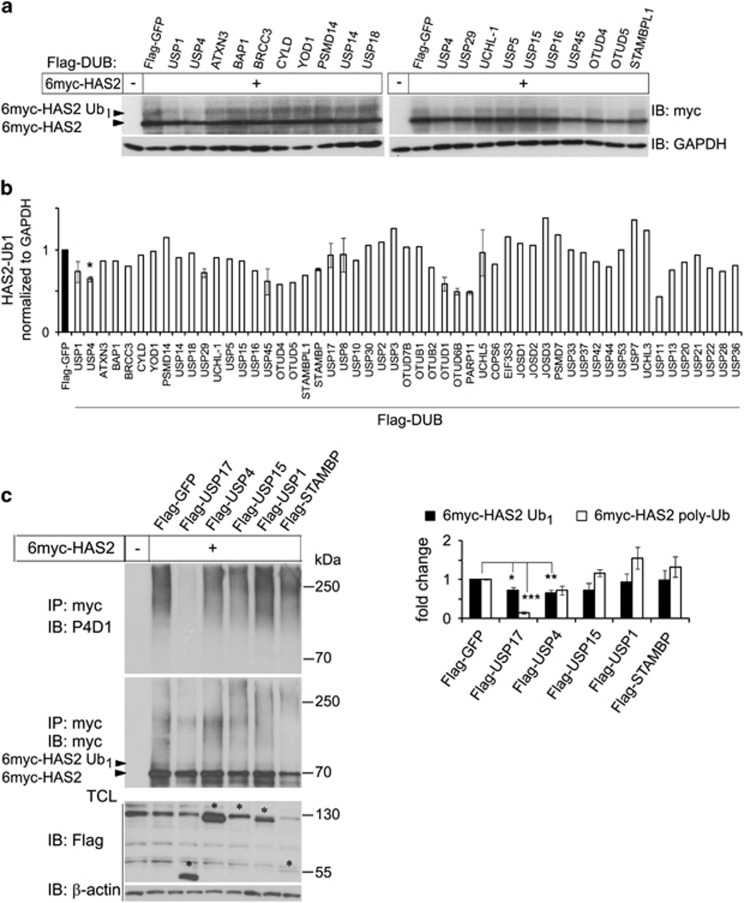
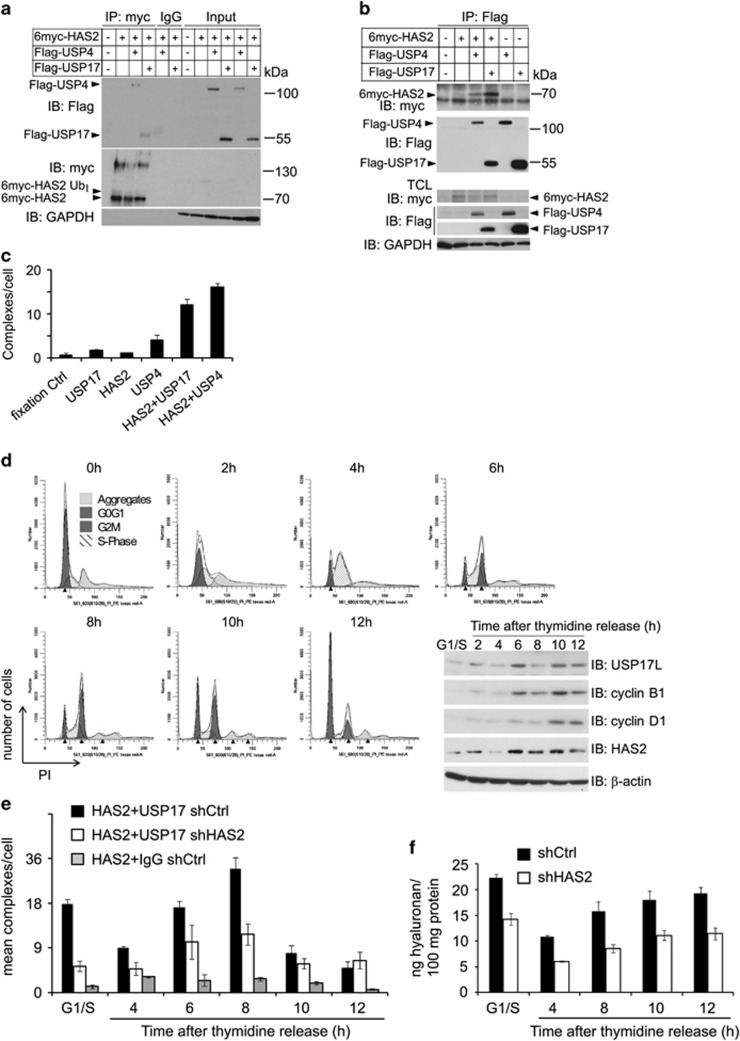
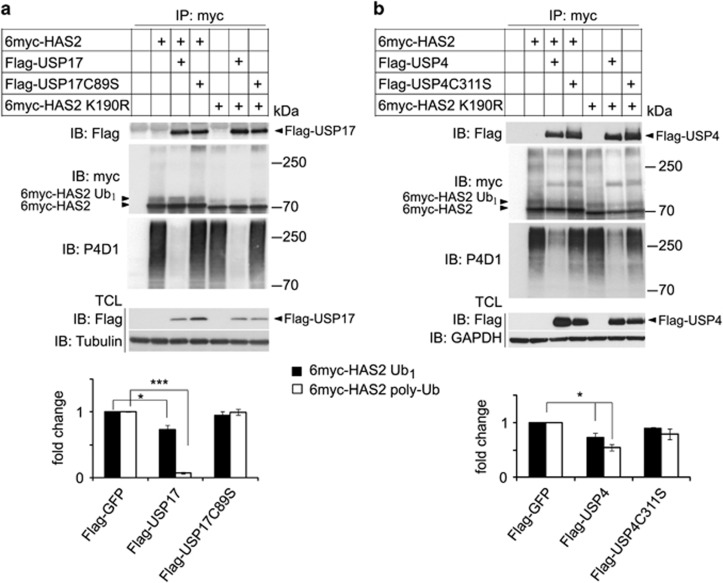
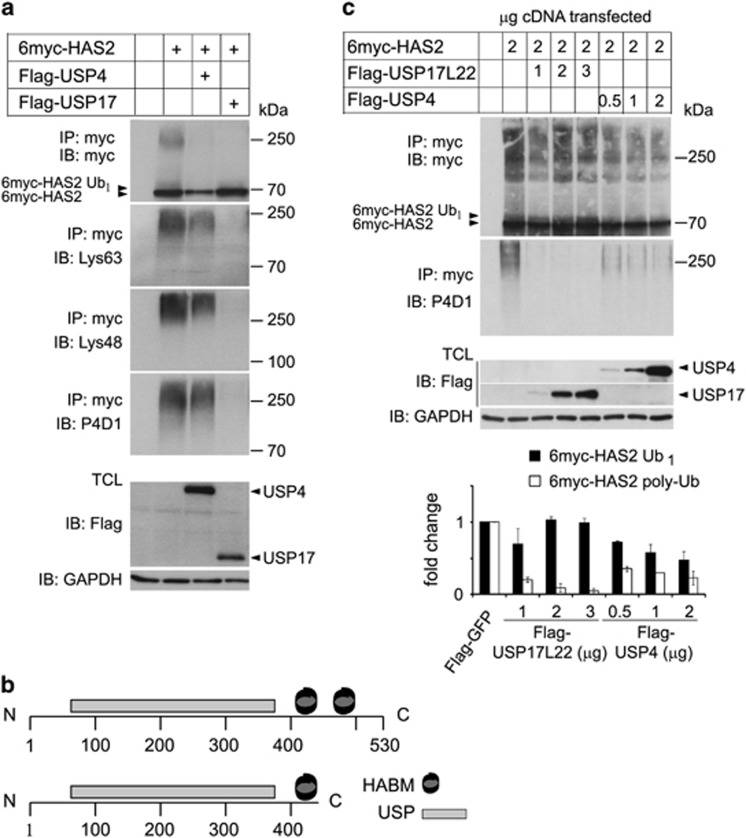
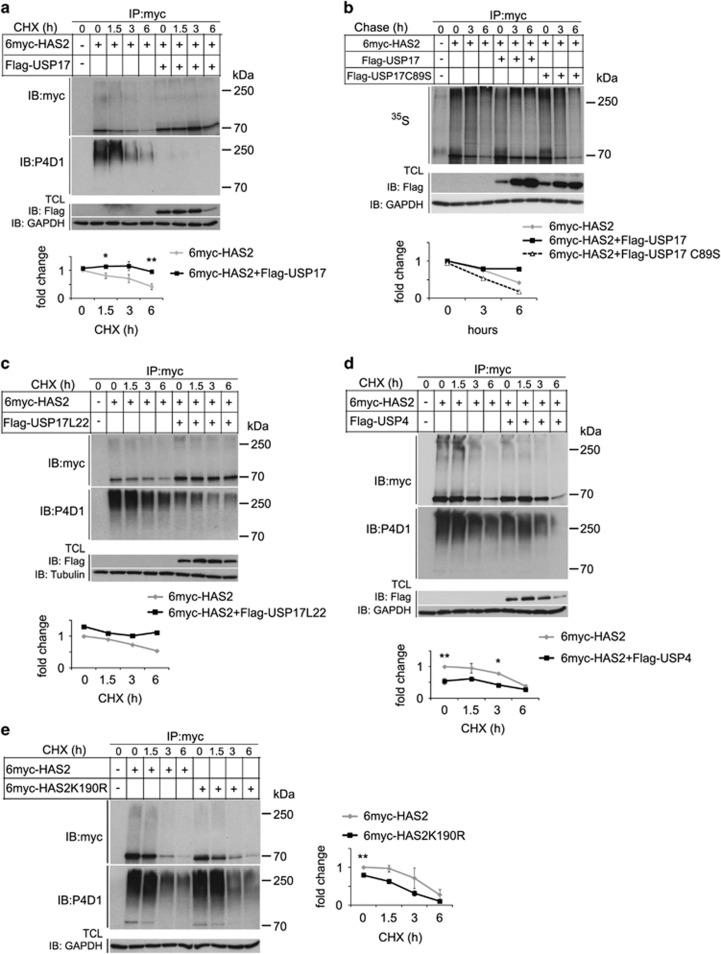
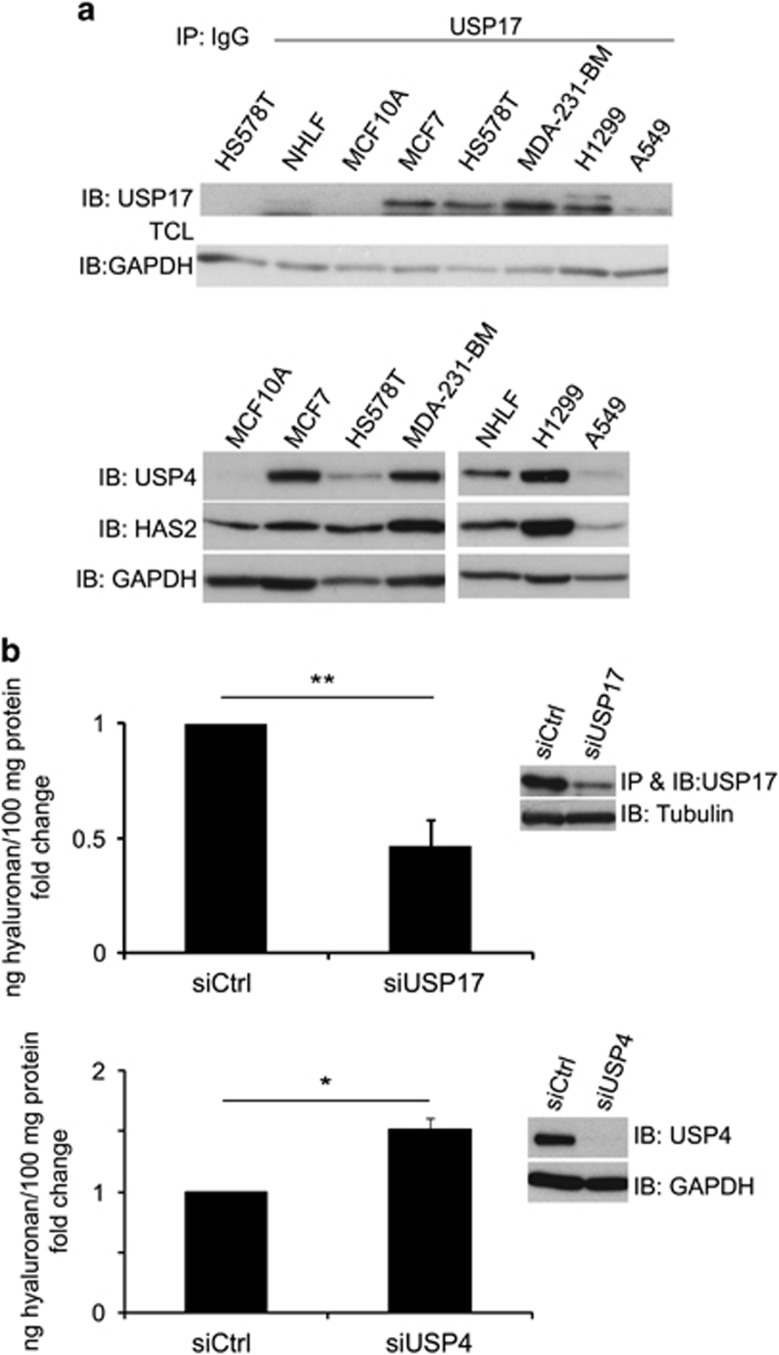
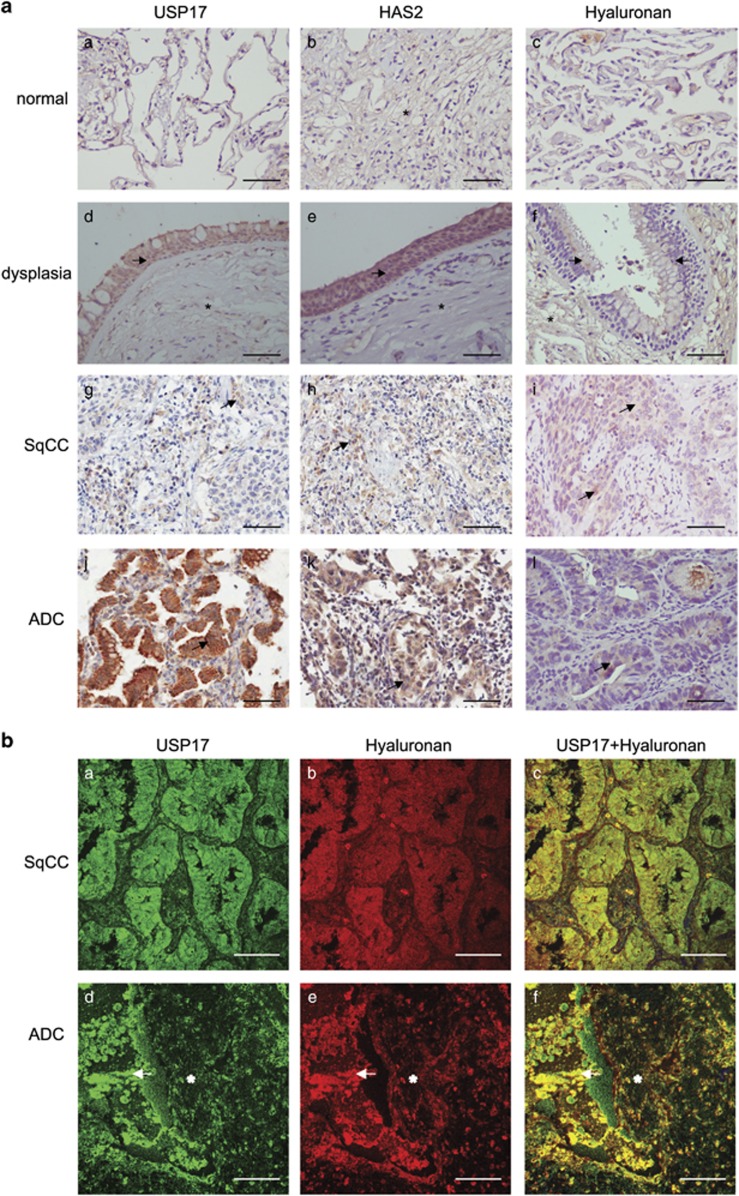
The levels of hyaluronan, a ubiquitous glycosaminoglycan prominent in the extracellular matrix, is balanced through the actions of hyaluronan-synthesizing enzymes (HAS1, 2 and 3) and degrading hyaluronidases (Hyal 1, 2, 3 and PH20). Hyaluronan accumulates in rapidly remodeling tissues, such as breast cancer, due to deregulated expression of the HAS2 gene and/or alterations of HAS2 activity. The activity of HAS2 is regulated by post-translational modifications, including ubiquitination. In order to identify deubiquitinating enzymes (DUBs) that are involved in de-ubiquitination of HAS2, a complementary (cDNA) library of 69 Flag-HA-tagged human DUBs cloned into retroviral vectors was screened in human embryonic kidney (HEK) 293T cells for their ability to de-ubiquitinate myc-tagged HAS2. Several DUBs were found to decrease the ubiquitination of 6myc-HAS2, among which, the most effective were USP17 and USP4. USP17 efficiently removed polyubiquitination, whereas USP4 preferentially removed monoubiquitination of 6myc-HAS2. Co-immunoprecipitation studies revealed interactions between HAS2 and USP17, as well as between HAS2 and USP4, in membrane preparations of HEK293T cells. USP17 significantly stabilized 6myc-HAS2 protein levels, whereas USP4 did not. The silencing of USP17 led to decreased hyaluronan production, whereas the suppression of USP4 increased hyaluronan synthesis. Importantly, high levels of USP17 and HAS2 were detected in a panel of cancer cell lines compared to normal cells, and immunohistochemical stainings revealed higher expression of USP17 and HAS2 in tissues of lung cancer patients compared to normal tissue. In conclusion, USP17 and USP4 differently affect HAS2 ubiquitination, and the stability and function of HAS2.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Watanabe K, Yamaguchi Y. Molecular identification of a putative human hyaluronan synthase. J Biol Chem 1996; 271: 22945–22948. - PubMed
-
- Shyjan AM, Heldin P, Butcher EC, Yoshino T, Briskin MJ. Functional cloning of the cDNA for a human hyaluronan synthase. J Biol Chem 1996; 271: 23395–23399. - PubMed
-
- Itano N, Kimata K. Molecular cloning of human hyaluronan synthase. Biochem Biophys Res Commun 1996; 222: 816–820. - PubMed
-
- Spicer AP, McDonald JA. Characterization and molecular evolution of a vertebrate hyaluronan synthase gene family. J Biol Chem 1998; 273: 1923–1932. - PubMed
-
- Spicer AP, Olson JS, McDonald JA. Molecular cloning and characterization of a cDNA encoding the third putative mammalian hyaluronan synthase. J Biol Chem 1997; 272: 8957–8961. - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
